2.1.1. Les organes d'osmorégulation
2.1.2. Animaux sténohalins et euryhalins
2.2.1. Les espèces isosmotiques
2.2.2. Les espèces hypoosmotiques
2.2.3. Les espèces hyperosmotiques
2.2.4. Les hyper - hypoosmorégulateurs
Comme nous venons de le signaler, les problèmes générés par l'existence de gradients osmotiques et ioniques entre les milieux extérieurs et intérieurs concernent essentiellement la régulation de pertes ou de gains d'eau et de pertes ou de gains de Na+ et Cl-. A ces problèmes doit s'ajouter celui du contrôle de la perte d'urée spécifique à Latimeria et aux chondrichthyens.
Les systèmes impliqués dans le maintien de ces gradients ainsi que dans la régulation de ces gains et pertes vont faire intervenir des phénomènes d'entrée et de sortie faisant eux-mêmes intervenir des mécanismes actifs et passifs. Les phénomènes passifs concernent les mouvements diffusionnels d'eau, de Na+ et Cl- ainsi que d'urée qui s'établissent suite à l'existence des gradients. Des phénomènes "actifs" consommant de l'énergie s'opposent à ces mouvements passifs. Ils interviennent dans le transport contre gradient des ions Na et Cl, le transport et la synthèse de l'urée ainsi que dans les mouvements osmotiques d'eau couplés à un transport actif d'ions.
2.1.1. Les organes d'osmorégulation
Les phénomènes impliqués, qu'ils soient actifs ou passifs, ne concernent finalement
qu'un nombre limité de structures (figure 1-9). Il y a tout d'abord
le tractus digestif qui peut être une source de gains d'eau et d'ions à un bout et une
source de perte d'eau et d'ions à l'autre. Il y a ensuite les systèmes dits rénaux
(néphridies de la plupart des invertébrés, tubules de Malpighi des insectes, néphrons
des vertébrés). Ces systèmes sont toujours une source de perte d'eau et d'ions. Il y a
aussi une série d'organes particuliers servant plus ou moins spécifiquement au transport
actif d'ions. Ils peuvent être selon le cas une source de gain ou de perte (voir plus
loin). Il s'agit des branchies des crustacés hyperosmorégulateurs et des poissons
téléostéens, de la peau des amphibiens, des glandes rectales des sélaciens ou encore
des glandes nasales, linguales ou sub-linguales des reptiles et des oiseaux. Ces glandes
sont encore appelées "glandes à sel" ou "organes extra-rénaux". Il
y a enfin les téguments externes qui sont le siège de l'essentiel des mouvements
diffusionnels. Ils peuvent être la source de pertes ou de gains d'eau et d'ions.
Figure 1-9: Les organes impliqués dans l'osmorégulation. OER: organes extra-rénaux. OR: organes respiratoires. SR: systèmes rénaux. TD: tube digestif. TE: téguments externes.
En envisageant plus particulièrement les mouvements diffusionnels, il faut faire un net distinguo entre les animaux ayant une perméabilité tégumentaire élevée et ceux ayant une perméabilité basse. Ceci est particulièrement vrai dans le cas de l'eau. Comme le montre la figure 1-10, le "turnover" de l'eau ramené à un poids unitaire (100 g) chez différentes espèces en conditions normales varie en gros de 0,05 à 0,5 ml par jour chez les espèces à perméabilité tégumentaire basse et de 10 à 70 ml par jour chez les autres: soit de 20 à plus de 1000 fois plus! Ces différences sensibles vont évidemment impliquer des activités compensatoires très différentes, beaucoup plus importantes chez les dernières espèces que chez les premières.
Figure 1-10: Comparaison du turnover de l'eau chez des espèces à perméabilité tégumentaire basse ou élevée.
Les différents mécanismes impliqués aux différents niveaux que nous venons d'évoquer rapidement seront donc mis en œuvre différemment suivant les espèces envisagées et leurs problèmes spécifiques. Dans ce cadre, il nous faut distinguer les animaux terrestres des animaux aquatiques et parmi ces derniers, les isosmotiques des hyperosmotiques et des hypoosmotiques. Tous ces animaux ont, comme le rappelle la figure1-8, des problèmes tout à fait différents.
2.1.2. Animaux sténohalins et euryhalins
Nous avons jusqu'à présent considéré des animaux vivants dans un milieu bien défini,
stable et n'en sortant pas. Les problèmes osmotiques et donc d'osmorégulation peuvent
devenir plus complexes chez les espèces aquatiques dites euryhalines, qui peuvent soit
changer de milieu comme certains poissons (anguilles, saumons) ou crustacés (crabe bleu,
crabe chinois), soit supporter des modifications de la salinité de leur milieu extérieur
(divers mollusques, vers, crustacés ou poissons vivant dans les régions littorales ou
d'estuaires). Chez ces espèces, l'osmolarité du milieu extérieur peut varier très
largement (entre l'eau douce et l'eau de mer et parfois l'eau de mer concentrée par
évaporation). Dans ces conditions, l'osmolarité sanguine peut évoluer de différentes
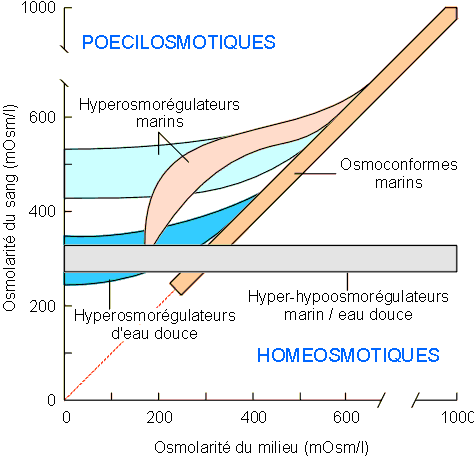
manières en fonction des espèces considérées (voir figure 1-11).
Dans ce cadre, nous devrons considérer des animaux dits osmoconformes,
hyperosmorégulateurs et hyper-hypoosmorégulateurs.
Figure 1-11: Evolution de l'osmolarité du sang en fonction de celle du milieu extérieur chez les espèces euryhalines.
Les osmoconformes sont tous marins. Chez eux le sang a toujours la même osmolarité que le milieu extérieur. Il s'agit des mollusques, de certains vers et crustacés. Leurs problèmes seront ceux des invertébrés isosmotiques.
Les hyperosmorégulateurs sont soit marins soit d'eau douce. Chez eux le sang peut être maintenu à une valeur supérieure à celle du milieu extérieur lorsque cette dernière est basse. En écophysiologie les hyperosmorégulateurs marins sont classiquement divisés en "forts" et "faibles" selon qu'ils peuvent s'adapter à l'eau douce ou qu'ils ne peuvent survivre qu'en eau saumâtre. Il s'agit de la plupart des crustacés marins et de certains vers marins. Tous les invertébrés d'eau douce sont hyperosmorégulateurs. Les hyper-hypoosmorégulateurs sont comme les précédents, soit marins soit d'eau douce. Chez eux le sang est maintenu à une osmolarité supérieure à celle du milieu extérieur lorsque cette dernière est basse; il est par contre maintenu à une osmolarité inférieure à celle du milieu extérieur lorsque cette dernière est élevée. On retrouve dans ce groupe tous les vertébrés aquatiques à partir des poissons téléostéens ainsi que quelques crustacés. Leurs problèmes seront ceux des hyperosmorégulateurs lorsqu'ils sont en milieu dilué et ceux des hypo-osmorégulateurs lorsqu'ils sont en milieu concentré. A la limite, l'osmolarité du sang peut rester constante quelles que soient les conditions extérieures. On parlera alors d'homéosmoticité. Les oiseaux et les mammifères sont homéosmotiques. Certains peuvent cependant ne pas l'être dans différentes conditions extrêmes (cfr. écophysiologie).
Nous allons donc envisager les mécanismes d'osmorégulation en fonction des problématiques particulières auxquelles les animaux ont à faire face. Dans ce cadre, nous distinguerons, comme nous l'avons dit, les animaux terrestres et les animaux aquatiques iso, hypo ou hyperosmotiques.
Comme nous l'avons signalé dans la section précédente, nous allons distinguer ici les animaux isosmotiques, ceux pouvant effectuer une hyperosmorégulation et ceux pouvant faire de l'hypoosmorégulation. Un résumé des mécanismes utilisés est présenté dans la figure 1-12, en fin de section 2.
2.2.1. Les espèces isosmotiques
Ces espèces sont toutes marines, elles peuvent être scindées en deux groupes. Le
premier comprend les agnathes et l'essentiel des invertébrés marins mis à part quelques
crustacés. Le second comprend les chondrichthyens, sélaciens et chimères, ainsi que Latimeria.
Les espèces du premier groupe n'ont guère de problèmes d'osmorégulation. Leur sang est en effet isosmotique à l'eau de mer extérieure et les effecteurs osmotiques majeurs sont les mêmes dans les deux milieux (Na+, Cl-).
Les espèces du second groupe n'auront guère de problèmes de balance hydrique, leur sang étant isosmotique au milieu extérieur. L'osmorégulation va porter ici sur une régulation de pertes diffusionnelles d'urée et de gains de NaCl.
Les pertes d'urée seront limitées par une réabsorption au niveau rénal. Elles seront compensées par une synthèse, via le cycle de l'urée, au niveau hépatique et rénal (voir plus loin: excrétion azotée). Les chondrichthyens paraissent être les seuls animaux capables d'effectuer une réabsorption rénale active d'urée importante. Le mécanisme impliqué reste à l'heure actuelle totalement inconnu. Il est très efficace. On a en effet pu démontrer que chez Squalus acanthias plus de 90 % de l'urée filtrée au niveau du rein est réabsorbée (urée filtrée: 1225 mmol/kg.h - urée excrétée: 115 mmol/kg.h).
Pour contrôler les gains de NaCl, ces espèces boiront peu, limitant ainsi une voie d'entrée importante. Un peu du NaCl en excès sera éliminé avec les urines qui sont isotoniques au sang. Ces mécanismes ne suffisent évidemment pas à contrebalancer les entrées par diffusion et l'excès de NaCl doit être éliminé par des organes sécréteurs "extra-rénaux". Chez les sélaciens il s'agit de la glande rectale, un organe situé au niveau du cloaque, à la fin du tractus digestif, qui peut produire un liquide hypertonique au sang. Les mécanismes d'élimination de l'excès de NaCl chez les chimères et les cœlacanthes sont à l'heure actuelle très peu connus.
2.2.2. Les espèces hypoosmotiques
On retrouvera dans ce groupe tous les vertébrés marins à partir des poissons
téléostéens ainsi que quelques crustacés marins. Comme les sélaciens , ces animaux
ont une concentration en NaCl sanguine largement inférieure à celle du milieu
extérieur. Il leur faudra donc limiter les gains diffusionnels et les contrebalancer par
excrétion active. De plus, l'osmolarité du sang est également largement inférieure à
celle du milieu extérieur. Il faudra donc combattre une tendance permanente à la
déshydratation. Cette tendance sera plus ou moins importante suivant les
caractéristiques de perméabilité des téguments extérieurs. Elle sera grande chez les
téléostéens et nettement plus faible chez les reptiles, oiseaux ou mammifères,
espèces revenues secondairement au milieu marin et ayant gardé les caractéristiques de
faible perméabilité à l'eau de leurs ancêtres terrestres. Chez la plupart de ces
espèces, la tendance à la perte d'eau sera compensée par la boisson. Certaines espèces
boivent donc beaucoup; c'est le cas des poissons téléostéens, chez d'autres espèces
cependant ou la perméabilité tégumentaire à l'eau est particulièrement faible, boire
n'est plus une nécessité et la prise de nourriture peut suffire à procurer l'eau
requise pour effectuer la balance hydrique. C'est semble-t-il le cas des mammifères
marins qui boivent très peu. Les lions de mer californiens en captivité peuvent par
exemple rester très longtemps (plusieurs semaines) sans boire, la balance hydrique étant
effectuée uniquement à partir de la nourriture qui leur est donnée.
L'eau soit bue, soit ingérée avec ou contenue dans la nourriture sera transférée vers le sang au niveau intestinal. Ce processus est consécutif à un transport actif de NaCl de la lumière intestinale vers le sang (voir plus loin et surtout livre II: physiologie cellulaire). Le NaCl impliqué dans l'entrée d'eau va donc s'ajouter aux gains par diffusion via les téguments. Un peu de ce NaCl sera éliminé avec les urines. Ces urines sont cependant peu abondantes pour ne pas aggraver la perte d'eau. Elles sont par ailleurs au mieux isotonique au milieu extérieur en NaCl, sauf chez les mammifères marins. L'élimination par cette voie ne sera donc pas suffisante, sauf chez les mammifères.
L'essentiel des hypoosmotiques marins va donc utiliser des glandes sécrétrices "extra-rénales" pour éliminer l'excès de NaCl. Comme chez les sélaciens, il s'agit d'organes pouvant réaliser un transport actif de NaCl contre gradient. Cette excrétion active de NaCl aura lieu au niveau des branchies chez les poissons et au niveau de glandes dites "à sel", linguales, sub-linguales ou nasales chez les reptiles et les oiseaux. Le tableau 1-3 reprend à titre d'exemple, un comparatif des productions urinaires et des glandes nasales chez un goéland pendant 175 minutes après l'ingestion d'eau de mer. On remarquera une augmentation de volume des deux côtés mais surtout une concentration en Na+ largement supérieure à celle du sang dans le fluide sécrété par la glande nasale. L'urine, elle, reste à une concentration en Na+ inférieure à celle du sang.
TABLEAU 1-3
Production de fluide au niveau du cloaque et des glandes nasales d'un goéland
pendant 175 min après ingestion de 134 ml d'eau de mer (± 1/10 du poids de l'animal)
| Glandes nasales | Cloaque | |||
| Temps (min) | Volume (ml) | Na+ (mM) | Volume (ml) | Na+ (mM) |
| 15 40 70 100 130 160 175 |
2,2 10,9 14,2 16,2 6,8 4,1 2,0 ------- 56,3 |
798 756 780 776 799 800 780 |
5,8 14,6 25,0 12,5 6,2 7,3 3,8 ------- 75,2 |
38 71 80 61 33 10 12 |
Chez les mammifères marins, c'est le rein qui va se charger de l'excrétion de l'excès de NaCl. Le rein de ces espèces, contrairement à celui de toutes les autres, est en effet capable de produire une urine très hypertonique au sang en NaCl. La baleine peut par exemple produire une urine dont la concentration en Cl- atteint plus de 800 mmol/l alors que le sang est à quelque 145 mmol/l. Chez les autres mammifères la capacité de concentration du rein en NaCl est nettement moins élevée, ce qui les rend incapables d'effectuer une balance hydrique positive à partir de l'ingestion d'eau de mer (tableau 1-4).
TABLEAU 1-4
Effet de l'ingestion d'un litre d'eau de mer sur la production d'urine chez l'homme et
la baleine
| Eau de mer consommée | Urine produite | Balance hydrique | |||
| Cl (mM) | Volume (ml) | Cl (mM) | Volume (ml) | (ml) | |
| Baleine Homme |
1000 1000 |
535 535 |
820 400 |
650 1350 |
+ 350 - 350 |
2.2.3. Les espèces hyperosmotiques
Ce groupe comprend toutes les espèces d'eau douce ainsi que la plupart des animaux
euryhalins (sauf osmoconformes) lorsqu'ils sont en milieu dilué. Les hyperosmotiques
auront à faire face à des gains d'eau et des pertes de NaCl.
Les entrées d'eau seront d'autant plus importantes que la perméabilité à l'eau des téguments est élevée. Les vertébrés à partir des reptiles ont conservé la perméabilité tégumentaire basse de leurs ancêtres terrestres. Leurs problèmes hydriques seront donc moins importants que ceux des invertébrés, des poissons et des amphibiens vivant en eau douce. Les espèces d'eau douce boivent donc en général très peu ou pas du tout de façon à limiter leurs entrées d'eau. Elles forment par ailleurs une urine abondante pour éliminer l'eau entrant essentiellement par diffusion via les surfaces tégumentaires.
En ce qui concerne les pertes diffusionnelles de NaCl, elles seront limitées par la production d'une urine très hypotonique au sang en NaCl. Tous les animaux d'eau douce sont capables d'effectuer une réabsorption de NaCl au niveau rénal. C'est également le cas des poissons euryhalins. Seuls les invertébrés marins euryhalins paraissent incapables de produire une urine peu concentrée en NaCl (cfr. tableau 1-5; C. maenas, C. magister, E. sinensis). Ces pertes seront compensées par les apports alimentaires lorsqu'elles ne sont pas trop importantes. C'est le cas chez les espèces ayant une perméabilité tégumentaire basse (reptiles, oiseaux, mammifères, insectes). Chez les crustacés ainsi que chez les poissons et les amphibiens, la perméabilité tégumentaire est élevée et les apports alimentaires ne suffisent pas. Ces espèces doivent en fait "pomper" activement des ions Na+ et Cl- dans le milieu extérieur pour assurer leur balance ionique. Ce sont les branchies qui effectuent ce travail chez les crustacés et les poissons. Chez les batraciens le transport d'ions est assuré par la peau. Les mécanismes intimes de ces systèmes de transport seront envisagés dans le livre II traitant de la physiologie cellulaire.
TABLEAU 1-5
Fonction urinaire chez différentes espèces hyperosmorégulatrices
| Habitat Milieu d'adaptation |
Production d'urine |
Na+ U/S efflux |
Osmolarité U/S |
||
| Crustacés euryhalins | |||||
| Carcinus maenas | EM 40% EM |
1,8 8,8 |
0,92 0,95 |
0,69 3,76 |
0,98 - |
| Cancer magister | EM 50% EM |
0,6 4,9 |
0,83 0,85 |
0,27 1,54 |
0,96 0,97 |
| Eriocheir sinensis | ED | 1,7 | 1,07 | 0,54 | 0,99 |
| Astacus fluviatilis | ED | 3,4 | 0,03 | 0,02 | - |
| Poissons euryhalins | |||||
| Anguilla anguilla | ED | 0,6 | 0,04 | 0,004 | - |
| Platichthys flesus | ED | 1,8 | 0,03 | 0,03 | 0,37 |
| Salmo gairdneri | ED | 4,7 | 0,07 | 0,05 | 0,15 |
| Carassius auratus | ED | 1,4 | 0,08 | 0,02 | 0,12 |
Un problème particulier concerne les protozoaires et les cœlentérés d'eau douce. Ces espèces sont capables de maintenir des concentrations tissulaires en Na, K et Cl supérieures à celles de leur milieu extérieur. On ne trouve cependant chez elles aucunes des structures impliquées chez les autres dans les mécanismes d'osmorégulation en milieu dilué.
Les protozoaires d'eau douce sont tous pourvus de vacuoles pulsatiles (une ou plusieurs) qui interviennent clairement dans l'élimination de l'eau qui envahit le cytoplasme par diffusion. Ces structures sont en général absente chez les espèces marines et parasites, isosmotiques à leur milieu extérieur. Le rythme de contraction des vacuoles est fonction de l'osmolarité du milieu; il décroît jusqu'à l'arrêt lorsque l'osmolarité extérieur atteint des valeurs de 90 à 120 mOsm/l, correspondant en gros à l'osmolarité moyenne du liquide intracellulaire. Le remplissage des vacuoles parait associé à un transport actif d'ions (Na+ et/ou Cl-); l'eau suit par mouvement osmotique. Le liquide vacuolaire est finalement expulsé par contraction. Chez de nombreux infusoires ciliés, la vacuole a un orifice permanent par lequel le liquide produit peut s'écouler en continu.
Les études concernant les cœlentérés restent à ce jour très peu nombreuses. Elle ne concernent de plus que deux espèces de cnidaires à des stades très différents: polype pour Hydra et méduse pour Craspedacusta. Ces organismes, comme tous les cœlentérés d'eau douce, sont animés de contractions rythmiques. La fréquence de ces contractions est d'autant plus basse que l'osmolarité du milieu extérieur augmente. Le mécanisme d'expulsion d'eau actuellement proposé fait dès lors intervenir un transport actif d'ions (Na et/ou Cl) des cellules de l'endothelium vers la cavité entérique; l'hyperosmolarité locale ainsi engendrée provoque un mouvement d'eau des cellules et de la mésoglée vers cette cavité. L'eau est ensuite expulsée par la bouche à la faveur des contractions.
2.2.4. Les hyper - hypoosmorégulateurs
Comme nous l'avons déjà signalé, certaines espèces euryhalines, essentiellement des
poissons mais aussi quelques crustacés, peuvent maintenir une osmolarité sanguine
supérieure à celle du milieu extérieur lorsqu'il est dilué et inférieure à celle-ci
lorsque le milieu est concentré (2.1.2.). Ces espèces auront donc
les problèmes des hyperosmorégulateurs en eau douce et ceux des hypoosmorégulateurs en
eau de mer. Les solutions apportées seront celles déjà décrites pour ces deux groupes.
Ainsi une anguille ou un saumon par exemple boiront nettement moins en eau douce qu'en eau
de mer. Ils produiront par contre une urine plus abondante en milieu dilué qu'en milieu
concentré. Les branchies seront impliquées soit dans une expulsion de NaCl en eau de mer
soit dans un pompage de NaCl en eau douce. Ce résultat n'est pas obtenu par inversion du
mécanisme de transport; l'ion activement transporté par les branchies des animaux en eau
douce est le Na+. Il s'agit du Cl- chez les animaux en eau de mer.
Lors du passage de l'eau de mer à l'eau douce, les mécanismes de transport du Na+
sont activés alors que ceux responsables du transport de Cl- sont inhibés.
L'inverse se produit lors de la migration inverse (cfr. Physiologie
cellulaire). Il semble également que, chez les poissons du moins, le passage en eau
douce puisse induire une diminution de perméabilité à l'eau et aux ions des surfaces
tégumentaires.
Les animaux terrestres auront à faire face à des pertes d'eau et de NaCl. Un des aspects essentiels de la vie terrestre est le danger de déshydratation. Deux embranchements animaux seulement ont réussi pleinement leur adaptation au milieu terrestre. Il s'agit des arthropodes et des vertébrés, à partir des reptiles. On rencontre bien quelques mollusques et même un amphibien dans des milieux désertiques mais ceux-ci représentent des exceptions. La plupart des invertébrés autres qu'arthropodes de même que les vertébrés inférieurs aux reptiles n'arrivent à se maintenir en milieu terrestre que par le choix d'un habitat suffisamment humide. Ces espèces ont en général une perméabilité tégumentaire élevée et donc peu de résistance à la perte d'eau.
La résistance à la perte d'eau est donc un facteur limitant de la pénétration des espèces en milieu terrestre. Pour un même type de tégument, l'évaporation sera d'une part fonction du rapport surface/volume de l'animal considéré. Dans les mêmes conditions, un petit animal perdra plus d'eau qu'un gros par unité de poids. Les pertes d'eau varieront d'autre part considérablement avec les conditions extérieures (température, degré hygrométrique, vitesse du vent). L'importance de ces paramètres fait que, chez les animaux à perméabilité élevée, le comportement intervient de façon souvent prépondérante dans la survie en milieu terrestre. Chez les animaux bien adaptés, la perméabilité à l'eau des surfaces tégumentaires est en général très basse, de l'ordre de 50 à 100 fois inférieure à celle des autres (tableau 1-6), les pertes par évaporation se situant entre 0,5 et 50 mg H20/h.cm2 pour une tension de vapeur de 1 mm Hg. Ces espèces pourront donc rester dans un air sec et chaud plus longtemps que les autres. Les adaptations comportementales n'en sont pas moins importantes pour autant. Ces problèmes sortent du cadre de ce chapitre. Ils seront discutés dans la partie consacrée à l'écophysiologie.
TABLEAU 1-6
Evaporation tégumentaire chez différentes espèces
| Espèce | Evaporation | Espèce | Evaporation |
| Ver de terre Grenouille Salamandre Escargot actif |
400 300 600 870 |
Rat Iguane Vers de farine Escargot inactif |
46 10 6 39 |
Les pertes d'eau auront lieu via les surfaces tégumentaires, les urines et les fèces ainsi que via des sécrétions spécialisées (glandes sudoripares, lacrymales, etc...) ou encore via les surfaces respiratoires (les échanges gazeux se faisant toujours en milieu aqueux, l'air au niveau des épithélia respiratoires ainsi que l'air exhalé sont saturés en eau - cfr. chapitre 3). La perte via les surfaces respiratoires et tégumentaires représente en gros de 60 à 80 % des pertes totales, les pertes respiratoires intervenant dans ce total pour 20 à 40 %. Les pertes via les fèces et les urines ne représentent donc en général que 20 à 40 % des pertes totales. Les animaux terrestres auront en général des fèces peu hydratés et une urine peu abondante, très concentrée. Chez certaines espèces, le type d'excrétion azotée évolue, passant de l'ammoniaque à l'urée ou à l'acide urique en fonction de la disponibilité en eau, pour permettre une concentration maximale des urines. Certains insectes peuvent ainsi produire des excreta plus secs que l'air dans lequel ils vivent. Tous les animaux terrestres seront en fait caractérisés par une capacité importante de récupération d'eau au niveau rénal (voir plus loin).
Les pertes de NaCl auront lieu via l'excrétion. Chez certaines espèces, des glandes spécialisées vont intervenir de façon non négligeable dans ces pertes. C'est par exemple le cas des glandes sudoripares chez les animaux utilisant la transpiration comme moyen de thermorégulation (cfr. écophysiologie, chapitre 12). Ces pertes sont largement limitées par la capacité, généralisée chez les animaux terrestres, à former une urine largement hypotonique au sang en NaCl.
Les gains en eau et en NaCl seront assurés essentiellement par l'alimentation (boisson et nourriture). Certains arthropodes disposent par ailleurs de mécanismes particuliers leur permettant d'absorber la vapeur d'eau de l'air ambiant lorsque l'humidité relative est de 70 % ou plus. Ces mécanismes restent à l'heure actuelle assez mal connus. Chez Arenivaga sp., un crabe du désert, ils paraissent associés à deux structures de type vésical présentes dans l'hypopharynx. Dans d'autres cas le site d'absorption ne serait pas buccal mais rectal.
Chez certaines espèces les pertes d'eau sont suffisamment limitées pour qu'un accès à l'eau libre (boisson) ne soit pas indispensable à la réalisation de la balance hydrique. Des rats kangourou, Dipodomys spectabilis, ont par exemple été maintenus plus d'un mois sans accès à l'eau avec pour toute alimentation des graines de céréales ne contenant que 6 % d'eau. L'essentiel de la balance hydrique est assuré dans ces conditions par de l'eau métabolique provenant de l'oxydation des métabolites de la graine. Sur quelques 4 semaines à 15 % et à 20 % d'humidité relative, un rat kangourou d'environ 75 g va perdre 60 ml d'eau dont 44 par évaporation, 2,5 via les fèces et 13,5 via les urines. Il mangera 100 g de grains qui lui amèneront 6 ml d'eau absorbée et 54 ml d'eau métabolique (d'après Schmidt-Nielsen, 1983).
Dans bien des cas, l'eau n'est pas toujours accessible librement. Certaines espèces
n'auront accès à l'eau que périodiquement (climats désertiques). Bon nombre d'animaux
terrestres, invertébrés comme vertébrés, font montre dans ce cadre, d'une certaine
capacité à supporter des variations non négligeables d'osmolarité sanguine et donc à
survivre à une certaine déshydratation. Nombre d'invertébrés, crabes et insectes,
pourront ainsi supporter des pertes de 20 à 50 % de leur eau corporelle, parfois plus,
sans problème apparent. Il en va de même chez les reptiles et amphibiens qui pourront
aller ainsi jusqu'à 60 % de perte. Même chez les mammifères, classiquement considérés
comme homéosmotiques, certaines espèces désertiques (chèvres, chameaux) pourront
supporter des pertes de 20 à 35 % de leur eau corporelle. Dans ces situations, comme dans
bien d'autres, la survie devient essentiellement dépendante de la capacité des cellules
à supporter des modifications d'osmolarité de leur milieu environnant.