3.3.1. Digestion de la cire d'abeille
3.3.2. Digestion de la cellulose
a. chez les invertébrés
b. chez les vertébrés1. Stockage gastrique et rumination
2. La coprophagie
Le tube digestif des eukaryotes abrite généralement des micro-organismes qui vont contribuer de façon plus ou moins significative, par leurs activités métaboliques, ŕ la fourniture de nutriments directement utilisables. Ainsi par exemple, on peut dénombrer dans les fčces chez l'homme plus de 400 espčces de bactéries appartenant ŕ dix genres différents. Dans de nombreux cas, la participation des micro-organismes ŕ la nutrition n'est pas fortuite mais s'inscrit dans le cadre d'une organisation symbiotique spécifique. Dans ces conditions, les apports sont loin d'ętre négligeables; souvent męme, ils sont essentiels ŕ la survie de l'hôte.
Les micro-organismes symbiotiques seront, selon les cas, intracellulaires ou extracellulaires, logés dans différentes parties du tube digestif ou dans des cavités cœlomiques.
Différents invertébrés, allant des cœlentérés aux mollusques, hébergent dans certaines cellules des algues vertes (zoochorelles) ou brunes (zooxanthelles). A la lumičre, ces algues produisent ŕ partir d'unités C-1 fournies par l'hôte, des sucres simples dont il bénéficie (tableau 5-2). Le bénéfice peut ętre substantiel. On a ainsi pu calculer que l'hydre verte détourne ŕ son profit prčs de 50 % du carbone fixé par photosynthčse par l'algue qu'elle héberge. Chez les cœlentérés marins, il semblerait męme que la production carbonée des algues symbiotiques puisse couvrir l’essentiel des besoins du métabolisme de base (Muscatine, 1990).
TABLEAU 5-2
Algues symbiotiques intracellulaires et produits fournis ŕ l'hôte
| Algue | Hôte | Composé fourni | |
| Zoochorelle | Paramecium bursaria | Protozoaire | Maltose |
| Spongilla lacustris | Glucose | ||
| Chlorohydra viridissima | Cœlentéré | Maltose | |
| Convoluta roscoffensis | Plathelminthe | Glucose | |
| Placobranchus ianthobapsus | Mollusque | inconnu | |
| Tridacna crispata | inconnu | ||
| Zooxanthelle | Aiptasia pulchella | Cœlentéré | Glycérol |
| Anthopleura elegantissima | Glycérol | ||
| Fungia scrutaria | Glycérol | ||
| Pocillopora damicornis | Glycérol | ||
| Zoanthus confertus | Glycérol | ||
| Tridacna crocea | Mollusque | Glycérol |
On peut inscrire dans un cadre semblable le cas de certains invertébrés de grandes
profondeurs vivant prčs des cassures tectoniques (rifts). La découverte de ces espčces
est assez récente, datant des années 80. Les bactéries impliquées ici sont capables
d'utiliser l'énergie fournie par oxydation du H2S et des sulfates qui abondent
dans les sources d'eau chaude fréquentes au niveau de ces cassures. L'énergie
d'oxydation obtenue sert ŕ phosphoryler de l'ADP en ATP qui lui-męme interviendra dans
la synthčse de différents composés ŕ partir d'unités C-1. Le cas le mieux étudié ŕ
l'heure actuelle est celui du pogonophore des Galapagos (Riftia pachyptila) qui
peut atteindre des dimensions respectables de plus d'un mčtre ! Ce pogonophore se
développe avec toute une faune extręmement particuličre basée sur la production
bactérienne importante au niveau des sources sulfureuses sous-marines. Chez cette
espčce, la cavité cœlomique est presque entičrement occupée par de larges poches
(trophosomes) bourrées de bactéries faisant de l'ATP et fixant du CO2.
Les micro-organismes se trouvent dans ce cas ŕ différents niveaux du tube digestif soit estomac ou poches stomacales, soit intestin ou cćca intestinaux. Ils participent ici ŕ la digestion des aliments ingérés, fournissant ŕ l'hôte des produits de dégradation de ces aliments (figure 5-14). Par ailleurs, ils servent eux-męmes de source alimentaire. Dans certains cas, la nutrition repose exclusivement ou presque exclusivement sur l'activité des micro-organismes hébergés, l'hôte n'ayant pas les enzymes digestives adaptées ŕ sa nourriture essentielle ou n'en ayant qu'une partie ou encore n'en disposant qu'en quantité insuffisante. Le rôle des symbiotes du tube digestif est parfaitement illustré par deux exemples : celui de la digestion de la cire d'abeille par les oiseaux de la famille des indicatoridć et celui de la digestion de la cellulose par les espčces végétariennes et xylophages.
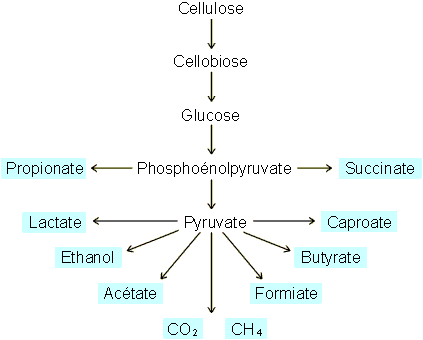
Figure 5-14: La dégradation anaérobie de la cellulose et ses produits terminaux par les micro-organismes du tube digestif de différents herbivores.
3.3.1. Digestion de la cire d'abeille
Le premier animal reconnu pour se nourrir de la cire des nids d'abeille fut la
larve du papillon Galleria mellonella, dans les années 1920. Curieusement, on ne
sait toujours pas exactement ŕ l'heure actuelle quels composants de la cire la chenille
utilise et comment elle les digčre.
La situation est plus claire en ce qui concerne les oiseaux africains formant le groupe des indicatoridć. Ces oiseaux sont connus pour se nourrir de la cire des nids d'abeille vers lesquels ils guident les personnes qui les suivent, d'oů leur nom anglais de "honey guide". La cire est chez eux digérée par des micro-organismes vivant dans l'intestin. Il s'agit pour l'essentiel d'une bactérie (Micrococcus cerolyticus) et d'une levure (Candida albicans). On a pu nourrir des poulets de cire d'abeille aprčs avoir inoculer dans leur tube digestif les micro-organismes qu'héberge l'intestin des indicatoridć.
3.3.2. Digestion de la cellulose
La cellulose est le polysaccharide de structure majeur des parois cellulaires
végétales. C'est donc un constituant important de l'alimentation chez de trčs
nombreuses espčces, tout particuličrement les herbivores et les xylophages. Chez ces
espčces, différents micro-organismes vont intervenir, de façon plus ou moins
importante, dans la digestion de la cellulose. Ils sont essentiellement anaérobies et
fournissent ŕ l'hôte des acides, produits du métabolisme oxydatif anaérobie du glucose
provenant de l'hydrolyse de la cellulose (figure 5-14). Il y aura
également production de gaz, essentiellement méthane et CO2, le méthane
paraissant provenir largement de la réduction du CO2.
a. chez les invertébrés
Dans la plupart des groupes, on trouve des cellulases endogčnes dont l'activité est
souvent assez faible et ne permet qu'une digestion trčs partielle de la cellulose. On
estime ainsi par exemple que le ver ŕ soie ne digčre que quelque 25 % de la cellulose
qu'il ingčre. Ce n'est cependant pas toujours le cas. Ainsi, chez Cténolepisma
lineata, un lépisme se nourrissant de feuilles d'eucalyptus, on estime que prčs de
80 % de la cellulose ingérée au cours d'un repas sont digérés par des cellulases
uniquement endogčnes. En général cependant, la plupart des espčces digérant bien la
cellulose hébergent des micro-organismes dont la contribution est importante. Chez les
insectes xylophages, il s'agit essentiellement de flagellés que l'on peut souvent
transplanter d'une espčce ŕ l'autre avec succčs. Ainsi par exemple le blattidć
xylophage Cryptocercus punctulatus contient au niveau du proctodeum des flagellés
symbiotiques qui assurent la digestion de l'essentiel de la cellulose ingérée. Ces
protozoaires peuvent ętre inoculés dans le tube digestif du termite Zootermopsis sp.
sans aucun problčme. Chez les termites inférieurs, généralement xylophages, on a
dénombré quelque 45 espčces différentes de flagellés symbiotiques, l'espčce la plus
généralement répandue étant Trichomitopsis termopsis. Chez ces termites, la
contribution des micro-organismes ŕ la digestion de la cellulose est essentielle; ils
meurent de faim une fois la faune intestinale tuée. De nombreux termites supérieurs ne
sont pas xylophages mais herbivores. Chez eux, la participation de cellulases exogčnes
est moins importante et on ne trouve que peu de flagellés symbiotiques. L'activité
cellulolytique endogčne est en général assez élevée pour digérer une partie
suffisante de la cellulose ingérée. Certaines espčces comme Macrotermes natalensis
trouvent par ailleurs des cellulases exogčnes dans une variété de champignon qu'elles
cultivent.
b. chez les vertébrés
Les vertébrés paraissent incapables de produire des cellulases. Les micro-organismes
cellulolytiques apparaissent donc comme un élément essentiel du systčme de digestion de
la cellulose chez tous les vertébrés et tout particuličrement chez les herbivores.
On trouve en fait des bactéries et des protozoaires symbiotiques produisant des cellulases pratiquement dans toutes les parties du tube digestif. Leur concentration ne devient cependant importante et donc significative dans le cadre de la digestion, que dans l'estomac et dans l'intestin. Dans la plupart des cas, les micro-organismes se trouvent en abondance uniquement dans la lumičre intestinale ainsi que dans des cćca et diverticules de la partie postérieure de l'intestin. Cette disposition prévaut des poissons aux oiseaux ainsi que chez de nombreux mammifčres, y compris l'homme et les primates. La localisation postérieure du site de digestion et la vitesse du transit intestinal diminue souvent largement le rendement de la digestion.
Chez différentes espčces, le systčme est rendu plus efficace par recyclage; les fčces sont en fait réabsorbées, ce qui permet de soumettre les matičres mal digérées ŕ une nouvelle attaque bactérienne. La coprophagie est ainsi largement pratiquée depuis les poissons jusqu'aux mammifčres. Chez différents mammifčres, le rendement est amélioré en reportant la digestion plus haut dans le tube digestif. Chez ces espčces, l'estomac et/ou l'œsophage présentent de larges poches bourrées de micro-organismes cellulolytiques. Ces poches permettent par ailleurs un stockage de longue durée, ce qui augmente encore le rendement de la digestion. Chez certaines espčces telles que les bovidés, le rendement est encore amélioré par rumination.
1. Stockage gastrique et rumination
La digestion de la cellulose avec stockage gastrique paraît limitée ŕ certains
mammifčres herbivores : tous les ruminants, certains singes, les kangourous, les
paresseux, les dugongs, l'hippopotame ainsi que certains rongeurs comme le hamster ou le
rat de Gambie. Chez toutes ces espčces, l'estomac est volumineux avec une partie
antérieure compartimentée en poches bourrées de bactéries, de phycomycčtes et de
protozoaires qui peuvent digérer la cellulose et d'autres matériaux végétaux qui s'y
accumulent. Le poids du contenu stomacal est en général considérable, variant le plus
souvent entre 10 et 25 % du poids de l'animal.
Chez les ruminants (chameaux, cerfs, bovidés, antilopes, moutons, chčvres, girafes), la digestion des matičres végétales est aidée par une régurgitation périodique du contenu de la partie antérieure de l'estomac; ce qui permet aux animaux de mâcher une nouvelle fois les fibres végétales pour les rendre plus accessibles aux attaques des micro-organismes.
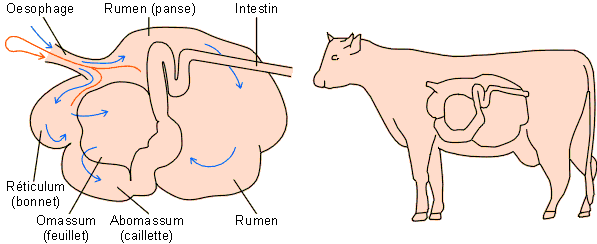
L'estomac des ruminants se subdivise en deux compartiments principaux séparés par une constriction transversale : l'estomac antérieur qui n'est en fait constitué que de chambres d'origine œsophagienne et l'estomac postérieur, seul glandulaire, comparable ŕ l'estomac classique des mammifčres. On compte ainsi un total de 4 poches ( figure 5-15).
Le rumen (panse) est la poche la plus volumineuse, représentant ± 80 % du volume total de l'estomac qui est de ± 250 l chez la vache.
Le reticulum (bonnet) est la plus antérieure et la plus petite, ne représentant que ± 5 % du volume gastrique total (± 15 l chez la vache).
L'omassum (feuillet), troisičme poche œsophagienne d'un volume de ± 8 % du volume total de l'estomac (± 20 l chez la vache).
L'abomassum (caillette) représente l'estomac classique des autres mammifčres. Son volume est égal ŕ celui de l'omassum (± 20 l chez la vache). C'est ŕ ce niveau qu'a lieu la sécrétion de HCl et de pepsinogčne et la premičre attaque acide des protéines.
Figure 5-15: L'estomac d'un ruminant (la vache) avec 3 poches d'origine œsophagienne (panse, bonnet, feuillet) et une poche (caillette) représentant l'estomac classique des mammifčres. Remarquez la place importante occupée par l'estomac dans l'animal.
Le rumen et le reticulum renferment une véritable culture de micro-organismes anaérobies parmi lesquels de nombreuses bactéries (tableau 5-3), des champignons inférieurs et des protozoaires ciliés appartenant ŕ deux familles essentiellement : les isotrichidć et les orphryoscolecidć. Leur concentration varie entre 105 et 1010 individus/ml (soit ± 2 ŕ 4 kg chez la vache par exemple). Comme l'indique le tableau 5-3 et la figure 5-14, les micro-organismes peuvent digérer de nombreux produits végétaux et produire, suite ŕ fermentation, une série d'acides gras volatils, du méthane et du CO2 en proportions variables en fonction du type de nourriture et des types de micro-organismes prédominants. En général, chez la vache, la concentration en acides gras volatiles atteint 100 ŕ 150 mmoles/l alors que la production de CH4 et de CO2 peut atteindre 1500 ŕ 2000 l/jour. L'essentiel du méthane et du CO2 sont éliminés par éructation; ce qui aide ŕ la rumination. Une bonne part des acides gras volatiles est absorbée directement au niveau des poches œsophagiennes dont le pH est tamponné ŕ des valeurs voisines de la neutralité par une abondante production de salive (200 l/jour chez la vache) riche en bicarbonates (pH 8,5). Le reste passe dans l'abomassum puis, aprčs attaque acide des protéines, passe dans l'intestin au niveau duquel ont lieu les autres séquences de la digestion, comme chez tous les mammifčres.
TABLEAU 5-3
Quelques bactéries de l'estomac des ruminants et leurs fonctions
| Espčces | Fonctions |
| Bacteroides succinogenes | C, A |
| Ruminococcus albus | C, X |
| Ruminococcus flavefaciens | C, X |
| Butyrivibrio fibrisolvens | C, X, PR |
| Clostridium lochheadii | C, PR |
| Streptococcus bovis | A, SS, PR |
| Bacteroides amylophilus | A, P, PR |
| Bacteroides ruminicola | A, X, P, PR |
| Succinimonas amylolytica | A, D |
| Selenomonas ruminantium | A, SS, GU, LU, PR |
| Lachnospira multiparus | P, PR, A |
| Succinivibrio dextrinosolvens | P, D |
| Methanobrevibacter ruminantium | M, HU |
| Methanosarcina barken | M, HU |
| Spirochete sp. | P, SS |
| Megasphćra elsderii | SS, LU |
| Lactobacillus sp. | SS |
| Anćrovibrio lipolytica | L, GU |
| Eubacterium ruminantium | SS |
Il faut par ailleurs considérer que les micro-organismes passant dans l'abomassum constituent un appoint nutritionnel non négligeable pour les ruminants. Ils procurent en effet ŕ leur hôte bon nombre de composés azotés (protéines et autres) ainsi que différents composés essentiels qu'ils synthétisent. On estime ainsi que chez la vache ± 70 % des 2 ŕ 4 kg de micro-organismes sont digérés par jour, fournissant, entre autres, plus de 100 g de protéines. Si les micro-organismes fournissent ŕ l'hôte une quantité notable de composés azotés, l'hôte fournit en retour aux micro-organismes une bonne part des groupements NH2 nécessaires ŕ la synthčse de ces composés. La vache récupčre en fait sous forme d'urée une bonne part des groupements NH2 provenant de son propre métabolisme protéique (cfr. chapitre 6). Elle récupčre de męme l'ammoniac provenant de l'activité bactérienne sur le bol alimentaire. L'urée ainsi produite est injectée dans le tube digestif avec la salive. Une partie importante de l'azote est ainsi recyclée : ce qui permet aux ruminants d'assurer sans trop de problčmes une balance azotée correcte alors que leur régime alimentaire peut ętre pauvre en produits azotés.
2. La coprophagie
L'ingestion des fčces est d'importance au plan nutritionnel pour bon nombre
d'espčces, des poissons aux mammifčres. L'autocoprophagie est cependant relativement
rare. Dans la plupart des cas, les animaux mangent les fčces d'autres espčces
(herbivores mangeant les fčces de carnivores par exemple), ce qui leurs procurent des
nutriments particuliers. Ce type de coprophagie peut ętre observé chez des espčces
telles que chien, chat ou cheval lorsque le régime alimentaire est mal équilibré.
L'autocoprophagie est pratiquée par de nombreux rongeurs (lapin, castor, écureuil,
chinchilla) ainsi que par certains lémuriens (opossum et koala). Chez ces espčces,
essentiellement herbivores, la digestion de la cellulose a lieu dans l'intestin; une
situation anatomique moins avantageuse que celle des ruminants et qui donne un rendement
moins élevé, compensé par réingestion immédiate de fčces particuličres
(cœcotrophes). Ces fčces ont la męme composition que le bol alimentaire se trouvant
dans l'intestin et contiennent donc tous les micro-organismes s'y trouvant. Les
cœcotrophes sont toujours expulsées séparément des autres fčces et sont nettement
plus molles, ne subissant que peu de réabsorption d'eau. Elles sont avalées telles
quelles sans ętre mastiquées et stockées dans une poche antérieure (non acide) de
l'estomac (figure 5-16), oů les attaques des micro-organismes
peuvent continuer. Ce comportement permet une nette amélioration du rendement de la
digestion de la cellulose. Il permet par ailleurs un gain en composés azotés et en
certains nutriments essentiels que procurent les micro-organismes lorsqu'ils sont
eux-męmes digérés.