|
 |
A côté du couple actine - myosine, un autre couple de structures protéiques joue un rôle important dans différents types de mouvements. Il s'agit du couple tubuline - dynéine que l'on retrouve à la base des mouvements des cils et flagelles. La tubuline parait également jouer un rôle essentiel dans certains transports protoplasmiques ainsi que dans le mouvement des chromosomes ayant lieu lors de la division cellulaire. Envisageons brièvement ces différents systèmes.
Les cils et flagelles sont des structures que l'on retrouve pratiquement dans tous les groupes zoologiques, des cœlentérés aux mammifères. Leurs battements dans différents types de cavités provoquent des mouvements de liquide, de mucus ou encore de nourriture particulaire (système respiratoire ou néphridien, cils des branchies de mollusques, etc. figure 7-29). Leurs battements dans le milieu extérieur sont utilisés comme système de propulsion chez bon nombre d'unicellulaires, chez certains pluricellulaires primitifs ainsi qu'au niveau de certains types cellulaires (urnes phagocytaires, spermatozoïdes...).
Figure 7-29: Exemples de structures ciliaires. A: Epithélium respiratoire (d'après Cross et Mercer 1995, modifié). B: Branchie de bivalve. Dans l'épithélium respiratoire, les cils sont implantés individuellement. Dans la branchie les cils forment un tapis ondulant en vagues. Les cils paraissent ici en relation les uns avec les autres par leurs structures d'ancrage cytoplasmique.
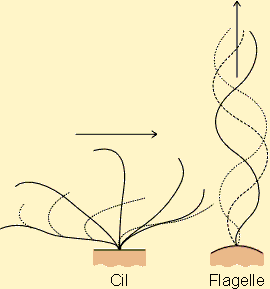
La distinction entre cils et flagelles se fait essentiellement sur base du type de mouvement que ces structures engendrent. Le flagelle provoque des mouvements de liquide parallèlement à son axe d'implantation. Ces mouvements seront par contre parallèles à la surface de la membrane dans le cas des cils (figure 7-30). Par ailleurs, les cils sont généralement présents en nombre, couvrant des surfaces importantes alors que les flagelles sont des structures uniques ou présentes en très petit nombre sur une cellule (deux ou trois).
La structure de base des cils et des flagelles est la même dans toute la série animale. Elle fait intervenir des architectures protéiques microtubulaires, elles-mêmes associées en structures plus complexes: axonème et cinétosome (voir plus loin). Les microtubules sont formés par association d'unités de tubuline a et b , deux molécules très semblables qui en s'empilant de façon alternée forment des protofilaments qui peuvent s'associer latéralement pour former des microtubules (figure 7-31).
Les microtubules peuvent exister sous forme stable, formant des structures permanentes; ce qui est le cas dans les cils et flagelles. Ils peuvent également se structurer à un bout pour se déstructurer à l'autre. Ce système association - dissociation peut devenir générateur d'un mouvement qui intervient dans différents types de transports protoplasmiques (cfr. 2.3 et 2.4 ci-après).
Figure 7-31: Formation des microtubules à partir d'unités de tubuline a et ß. Les unités de tubulines forment des hétérodimères qui peuvent s'empiler et s'associer latéralement pour se réunir en une structure cylindrique comprenant 13 protofilaments et ayant un diamètre extérieur de 28 nm avec un canal central de 14 nm. Généralement, il existe différentes protéines associées aux microtubules (PAMS) qui occupent les sillons existant entre protofilaments et hétérodimères.
Dans les cils et flagelles, les microtubules s'associent en une structure motrice centrale appelée axonème (figure 7-32). C'est un système très complexe auquel participent plus de deux cents protéines distinctes. Très brièvement, l'axonème est constitué d'un cercle de 9 doublets microtubulaires chacun comprenant en avant un microtubule A, complet (13 protofilaments) et en arrière un microtubule B, incomplet (10 protofilaments). Les différents doublets sont associés entre eux par des ponts de nexine. Les tubules A portent par ailleurs des bras de dynéine qui peuvent s'accrocher au tubule B du doublet précédent. Au centre de ce système se trouve une structure formée d'une gaine interne et de deux microtubules complets. Les microtubules A du système périphérique sont reliés à la gaine centrale par des fibres rayonnantes.
A l'endroit où le cil s'enfonce dans le cytoplasme, la structure de l'ensemble se modifie, formant un cinétosome ou corpuscule basal (figure 7-32). Les unités microtubulaires périphériques acquièrent un troisième microtubule incomplet et les bras de dynéine disparaissent. Les tubules centraux disparaissent également et des lamelles radiales relient les unités périphériques à la gaine centrale. Des protéines associées, partant vers l'extérieur assurent l'ancrage cytoplasmique. Le corpuscule basal peut, le cas échéant, donner naissance à un nouveau cil suite à amputation par exemple.
|
B |
Figure 7-32: A: Axonème, unité motrice de cils et flagelles et cinétosome d'ancrage. Explications dans le texte. B: Vue en microscopie électronique à transmission d'une coupe transversale dans les cils tapissant la cavité cœlomique de l'annélide Tomopteris. |
Les mouvements des cils et flagelles sont en fait liés à des ondulations de l'axonème dues à des glissements des doublets de microtubules les uns par rapport aux autres. Ces glissements sont induits par des mouvements d'avant en arrière des bras de dynéine. Un peu comme les filaments de myosine des muscles, les bras de dynéine sont formés d'une queue de laquelle émergent deux ou trois têtes globulaires dont l'angulation par rapport à la base commune peut varier. Le mouvement est lié à une hydrolyse d'ATP et les bras de dynéine présentent une activité ATPasique marquée. La force exercée par le mouvement d'un bras de dynéine a pu être estimée à 0,5 - 1 pN, une valeur proche de la force produite par le mouvement d'une tête de myosine. Les détails des interactions régissant les mouvements et l'accrochage des têtes globulaires des bras de dynéine aux microtubules sont toujours à l'étude actuellement. De même, la propagation des ondulations le long de l'axonème reste très mal comprise. Les mouvements de cils en vagues dites métachroniques (figure 7-33) restent également très mal compris. Il semble que les mouvements ondulatoires d'un cil puissent être initiés par une stimulation mécanique et soient indépendants de l'intégrité de la cellule sous-jacente. Dans ce cadre, une ondulation engendrée par le mouvement brusque d'un cinétosome pourrait se propager tout le long d'un cil et aux cils adjacents par couplage visco - mécanique impliquant les mouvements du milieu environnant.
Une partie du trafic intracellulaire de différentes molécules et architectures macromoléculaires paraît faire intervenir des structures microtubulaires. Ce type de mouvement est particulièrement étudié à l'heure actuelle au niveau des longs prolongements axonaux des cellules nerveuses. On a en effet pu mettre en évidence à ce niveau des mouvements relativement rapides (jusqu'à 40 cm par jour ! ) de vésicules et de matériel protéique le long d'axes microtubulaires. Ils pourraient faire intervenir un système d'association - dissociation des hétérodimères de tubulines générant le mouvement de différentes protéines associées telle la kinésine. Ces molécules serviraient de structures d'ancrage pour les matériaux à transporter (figure 7-34). Dans une autre hypothèse, la protéine d'ancrage pourrait voyager le long du microtubule servant alors en quelque sorte de rail de guidage (figure 7-34). Le fait que l'eau lourde qui stabilise les microtubules polymérisés, bloque les transports intracellulaires suggère que ces structures ne peuvent participer à la motilité cellulaire que si la possibilité de déplacer l'équilibre entre tubulines libres et polymérisées est maintenue. Le mécanisme effectif reste de toute façon très loin d'être élucidé.
Figure 7-34: Deux systèmes hypothétiques faisant intervenir les microtubules dans le transport rapide de matériel axonal. Dans un cas, le matériel s'accroche à une protéine qui s'intègre à un microtubule en extension. Dans l'autre, le matériel "glisse" le long d'un "rail" protéique intégré à un microtubule existant.
Si le clivage de la cellule paraît faire intervenir des filaments d'actine, il semble que le mouvement des chromosomes qui précède cette opération à l'anaphase fasse intervenir l'organisation d'un réseau de microtubules. Ceux-ci s'organisent en fait en un fuseau à partir du centrosome situé à un des pôles de la cellule. Lors de la prophase, les deux centrioles contenus dans le centrosome se dédoublent et migrent vers les pôles opposés de la cellule alors qu'un réseau de microtubules apparaît entre eux. Finalement, le réseau de microtubules, tendu entre les deux paires de centrioles occupant les pôles de la cellule, entoure le noyau dont l'enveloppe commence à se désorganiser. Les centrioles apparaissent alors comme de courts cylindres formés de neufs triplets de microtubules. Ces structures ressemblent beaucoup aux cinétosomes formant les racines des cils et flagelles (voir figure 7-32). A la métaphase, après désagrégation de la membrane nucléaire, les chromosomes migrent et s'attachent par leur kinétochore aux microtubules. Les kinétochores deviennent alors des centres organisateurs d'où croît une deuxième vague de microtubules, l'ensemble formant le fuseau le long duquel les chromatides sœurs résultant du dédoublement des chromosomes vont migrer au cours de l'anaphase (figure 7-35). Le mécanisme responsable de ce mouvement est loin d'être compris malgré les nombreuses recherches qu'il a suscitées, et suscite encore. Le mouvement pourrait être du à un phénomène d'association - dissociation des hétérodimères de tubulines. Il pourrait également faire intervenir un glissement de microtubules les uns par rapport aux autres un peu comme dans le battement des cils et flagelles. Enfin, il pourrait également mettre en œuvre des fibres d'actine. Dans ce dernier cas, les microtubules serviraient essentiellement de "rails" guidant les chromosomes. Le mouvement serait alors du à des interactions actine - myosine. Assez récemment, il a été mis en évidence une protéine, la dynamine, qui pourrait générer le glissement des microtubules du fuseau les uns par rapport aux autres; un peu comme le fait la dynéine dans les mouvements ciliaires. Le mécanisme finalement impliqué reste cependant totalement inconnu à ce jour.
Figure 7-35: Prophase, métaphase et anaphase: mouvement des centrioles, mouvement des chromosomes et accrochage au fuseau puis mouvement vers les pôles. Tous mouvements dont les mécanismes de base restent inconnus à ce jour (détails dans le texte).
De nombreuses espèces, invertébrés comme vertébrés, possèdent dans leurs téguments extérieurs des cellules pigmentées en forme d'étoile, appelées chromatophores. Ces cellules envoient des prolongements radiaires le long desquels des granules de pigments colorés paraissent se déplacer en direction centrifuge ou centripète parfois à une vitesse remarquable (cas des caméléons, des pieuvres ou des calmars par exemple).
Le mouvement dans les chromatophores tout comme celui des chromosomes reste à l'heure actuelle assez mystérieux. Il dépend clairement de l'organisation d'un réseau microtubulaire le long des axes duquel les pigments colorés se déplacent. La ou les protéines impliquées effectivement dans le déplacement restent inconnues (Fujii et Oshima, Adv. Comp. Environ. Physiol. 20, 1, 1994). Les mouvements sont essentiellement sous contrôle nerveux et hormonal. D'autres facteurs, notamment environnementaux, paraissent également pouvoir intervenir.