|
Figure 2-1: Les systèmes de propulsion des liquides circulants. A: Pompe péristaltique. B: Cœur à chambre. C: Pompe musculaire (veineuse). |
2.1.1. Les cœurs à chambres
2.2.1. Généralités
2.2.2. Commande et activité électrique - bruits du cœur
2.2.3. Relation entre activité cardiaque, masse corporelle et demande d'oxygène
La propulsion des liquides circulants peut faire intervenir différents systèmes ( figure 2-1).
|
Figure 2-1: Les systèmes de propulsion des liquides circulants. A: Pompe péristaltique. B: Cœur à chambre. C: Pompe musculaire (veineuse). |
1) Pompe péristaltique
Un premier système consiste en une sorte de pompe péristaltique dans laquelle une
portion d'un gros vaisseau est pourvue de structures musculaires. C'est l'alternance de
contractions et de relâchements qui induit le mouvement dans une direction donnée. On
retrouve ce système dans les cœurs tubulaires des tuniciers et des arthropodes (
figure 2-2) ainsi que dans les vaisseaux pulsatiles servant de cœur
chez les annélides et certaines holothuries mais aussi de système de propulsion annexe
dans les ailes des chauves-souris ou dans le conus arteriosus des élasmobranches par
exemple (figure 2-2).
2) Pompe musculaire
Un second système fait intervenir l'écrasement d'un vaisseau par des masses
musculaires en contraction. La présence de valvules assurent ici la direction de
circulation. On retrouve ce système de pompe musculaire dans les pompes veineuses
facilitant, chez l'homme par exemple, le retour du sang des jambes vers le cœur.
3) Cœur à chambres
Un troisième système fait intervenir une structure de propulsion spécifique dite
cœur à chambres. C'est de loin le système le plus répandu. Il consiste en une ou
plusieurs chambres entourées de muscles dont la contraction provoque un mouvement de
fluide. La direction est assurée par des valvules d'entrée et de sortie.
Figure 2-2: Différents types de cœur. A: Tubulaire (criquet). B, C: A une chambre (daphnie, homard). D: A deux chambres (escargot). E, F: Multichambres (téléostéen, requin). D'après Florey 1969, modifié (D); Hoar 1975, modifié (A, B, C); Schmidt-Nielsen 1983, modifié (E, F). Cœurs de reptiles: voir figures 2-6 et 2-7 . Cœur de mammifère: voir figure 2-8.
2.1.1. Les cœurs à chambres
Le modèle le plus primitif est un système à deux chambres tel qu'il en existe chez
différents invertébrés, mollusques notamment. Une chambre antérieure, l'atrium,
reçoit par un tronc veineux le sang de retour des tissus. Elle est en général peu
musclée. La chambre postérieure, le ventricule envoie par un tronc artériel, le sang
vers les tissus. Elle a une fonction essentiellement foulante et est munie d'une puissante
musculature, le myocarde. L'ensemble est généralement entouré d'un péricarde, membrane
épaisse et résistante jouant essentiellement un rôle de protection. Le péricarde, par
sa rigidité relative, peut par ailleurs aider au remplissage de l'oreillette lorsque le
ventricule se contracte. La puissance de contraction ventriculaire met en effet la cavité
péricardique en légère dépression. Ce système de base est extrêmement répandu. On
le retrouve, avec de notables modifications et adaptations, chez de nombreux
invertébrés, mollusques, crustacés, insectes, etc... ainsi que chez tous les
vertébrés. D'autres modèles sont apparus au cours de l'évolution mais ont eu beaucoup
moins de succès. Ainsi par exemple le cœur caudal des myxines qui fonctionne comme
une pompe aspirante et foulante (figure 2-3). Le jeu des muscles
modifie ici la position d'une structure cartilagineuse centrale en forme de T avec pour
conséquence l'accroissement du volume d'une chambre (aspiration) et la diminution du
volume de l'autre (pulsion). Ce type de cœur reste un cas particulier.
Chez les poissons, la structure cardiaque de base se complète d'un sinus veineux en avant de l'atrium qui reçoit le sang et d'un bulbe cardiaque à la suite du ventricule. Cette dernière structure peut être réduite voir absente chez certaines espèces. Elle porte le nom de cone artériel chez les sélaciens. Elle peut être suivie chez les téléostéens d'un bulbe artériel. Ces différentes structures précédant l'aorte sont munies de parois épaissies par la présence de fibres élastiques. Leur élasticité permet d'absorber les à-coups de pression dus au travail cardiaque, régularisant ainsi le flot sanguin.
Le sinus veineux disparaît avec le cloisonnement atrial chez les amphibiens. Le bulbe est maintenu chez les amphibiens et disparaît chez les reptiles. Sa fonction de régularisation du flot sanguin est reportée sur la première partie de l'aorte qui possède une épaisse paroi contenant des fibres élastiques.
L'évolution la plus marquante du système cardiaque est son cloisonnement aboutissant en fait chez les oiseaux et les mammifères ainsi que chez certains reptiles à deux structures juxtaposées mais complètement séparées (figure 2-4). Cette évolution transforme donc le cœur d'un système de propulsion essentiellement veineux chez les poissons, faisant circuler un sang désoxygéné venant des tissus vers les branchies en un système double, veineux et artériel, faisant circuler un sang désoxygéné des tissus vers les organes respiratoires d'une part et un sang oxygéné des organes respiratoires vers les tissus d'autre part. Cette évolution permet un accroissement d'efficacité considérable. Elle réalise en effet une relance du flot sanguin au sortir du système respiratoire. Ce dernier présente de fait des caractéristiques (organisation, fragilité) peu compatible avec le passage du flot sanguin à la pression qu'exige le maintien d'un débit important au niveau du reste de la masse tissulaire.
Figure 2-4: Cloisonnement du cœur et gain d'efficacité. Organisation d'une circulation systémique et d'une circulation pulmonaire. A: Poisson. B: Mammifère. A: atrium; OG, OD: oreillettes gauche et droite; V: ventricule; VG, VD: ventricules gauche et droit.
Le cloisonnement du cœur va en fait de pair avec l'individualisation d'une circulation respiratoire (petite circulation) et d'une circulation dite générale ou systémique (grande circulation), irriguant les autres organes. Cette disposition va permettre l'exploitation des deux systèmes circulatoires selon des modalités spécifiques. Chez les mammifères, par exemple, le circuit pulmonaire fonctionne à basse pression alors que le circuit systémique fonctionne à haute pression (cfr. tableau 2-1). Ce type d'organisation est déjà fonctionnel chez les varans malgré un cloisonnement incomplet (figure 2-7A).
TABLEAU 2-1
Paramètres cardiaques chez différentes espèces au repos
| Animal |
Pression artérielle (Sys/dia-mmHg) Systémique | Respiratoire |
Fréquence cardiaque (Puls/min) |
Volume d'éjection (ml) |
Débit cardiaque (ml/min) |
Débit/kg (ml/min.kg) |
|
| Mammifères | ||||||
| Homme Cheval Vache Rat Ecureuil Dauphin |
120/75 171/103 160/110 130/91 139/99 150/121 |
25/10 - - - - 55/35 |
75 48 50 360 276 100 |
77 1350 870 0,18 0,5 109 |
5775 64800 43500 65 138 10900 |
80-90 150 115 209 313 60 |
| Oiseaux | ||||||
| Pigeon Poulet Canard |
135/103 149/43 - |
- - - |
100 270 180 |
- 1,1 2,5 |
- 220 450 |
- 220 300 |
| Reptiles | ||||||
| Tortue Crocodile Iguane |
25/10 45/30 48/37 |
24/19 18/9 30/13 |
23 20 38 |
3,7 - 1,5 |
85 - 57 |
57 - 57 |
| Amphibiens | ||||||
| Grenouille Xenope |
35/21 32/21 |
31/16 31/11 |
40 43 |
0,035 0,056 |
1,4 2,4 |
20 24 |
| Poissons | ||||||
| Salmo gairdneri Pseudopleuronectes Squalus |
32/27 30/25 17/16 |
45/33 29/30 30/24 |
37,8 29,4 45 |
0,46 1,60 - |
17,8 47,0 - |
17,8 11,2 - |
| Mollusques | ||||||
| Octopus Loligo Helix Littorina |
28/10 56/21 21/5 3/1 |
5/2 - - - |
45 102 22 26 |
1,0 0,13 0,03 0,0025 |
45 13 0,6 0,065 |
45 13 60 6 |
| Crustacés | ||||||
| Cancer Carcinus Homarus Palinurus |
10/6 9/6 14/2 36/16 |
- - - - |
120 92 90 64 |
2,1 1,3 0,25 1,4 |
252 119 22,5 90 |
252 150 40 128 |
Le cloisonnement du cœur commence avec le cloisonnement de l'atrium en deux oreillettes. Cette étape est liée à l'apparition du système pulmonaire et à la différentiation de veines ramenant le sang oxygéné des poumons vers le cœur. Chez les dipneustes, les branches gauches et droites du système circulatoire pulmonaire fusionnent en une veine pulmonaire, qui amène le sang oxygéné à l'oreillette gauche. Le sang désoxygéné venant des tissus est lui ramené à l'oreillette droite par les veines caves. Le cloisonnement anatomique complet du ventricule et donc la séparation totale des deux circulations n'est effectuée que chez les reptiles crocodiliens, les oiseaux et les mammifères (figure 2-5).
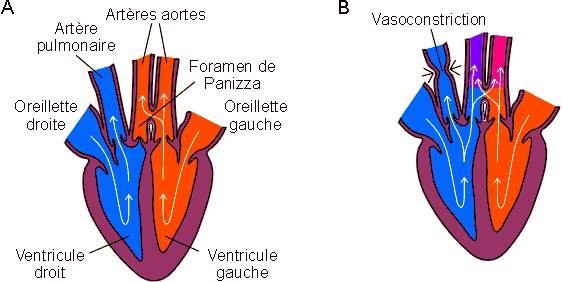
Chez les crocodiliens, le sang oxygéné du ventricule gauche peut cependant encore enrichir le sang désoxygéné venant du ventricule droit. Les artères provenant des deux ventricules restent en effet en connexion par le foramen de Panizza maintenu ouvert par des processus cartilagineux.
Figure 2-5: Les étapes du cloisonnement cardiaque. A: Poissons, sauf dipneustes. B: Dipneustes, amphibiens, reptiles non-crocodiliens. C: Reptiles crocodiliens. D: Oiseaux, mammifères. A: atrium; FP: foramen de Panizza; OG, OD: oreillettes gauche et droite; SR: système respiratoire; T: tissus; V: ventricule; VG, VD: ventricules gauche et droit.
L'organisation anatomique du système est cependant telle qu'en conditions normales, le sang désoxygéné paraît ne pas pénétrer de manière significative dans la circulation générale comme le montre des mesures de charge en oxygène du sang. En plongée par contre, la résistance du circuit pulmonaire augmente vraisemblablement par vasoconstriction contrôlée. L'augmentation de pression a pour résultat l'ouverture forcée du ventricule droit sur une artère aorte, avec recyclage d'une partie du sang entrant dans ce ventricule dans la circulation systémique (figure 2-6). L'avantage réel de ce système n'est pas clair. Il permet en tout cas d'éviter un engorgement du ventricule droit suite à la diminution de débit dans le circuit pulmonaire qui intervient en apnée (plongée).
Figure 2-6: Irrigation cardiaque chez les crocodiliens. A: Conditions normales. B: Plongée et vasoconstriction sur le circuit pulmonaire.
Chez les dipneustes, les amphibiens et les reptiles non-crocodiliens, le cloisonnement anatomique du ventricule n'est pas complet. Il existe cependant des septa réalisant un cloisonnement partiel. L'organisation de ces septa ainsi que celle de septa existant à d'autres niveaux réalisent semble-t-il un cloisonnement fonctionnel efficace. Les mesures montrent en effet qu'il y a peu de mélange des sangs oxygénés et désoxygénés. On estime en effet qu'une dizaine de % seulement du sang arrivant au ventricule participent à un échange dans les shunts gauche-droite ou droite-gauche que créent les septa incomplets (figures 2-7A et 2-7B). L'importance de ces shunts est probablement contrôlable, permettant d'adapter les disponibilités en oxygène aux situations générées par le type de respiration bimodale qu'utilisent nombre de ces espèces. Il s'agit notamment du passage d'une respiration aquatique branchiale ou cutanée à une respiration aérienne pulmonaire et vice versa. A notre connaissance ce problème est malheureusement resté très peu étudié jusqu'à présent.
Figure 2-7A: Distribution du flot sanguin chez les reptiles non crocodiliens. Anatomie cardiaque et évolution de la pression lors d'une contraction au niveau de différentes structures cardiaques. A: Chez la tortue, il n'y a que peu de mélange des sangs provenant des atria (oreillettes) gauche et droit bien que ceux-ci débouchent tous deux au dessus de la partie gauche du ventricule (cavum arteriosum et cavum venosum). Lors de la systole, le système fonctionne comme une pompe unique avec des pressions identiques au niveau des circuits pulmonaires et systémiques. B: Chez le varan, la réduction de volume du cavum venosum et l'organisation différente des septa permet une répartition encore meilleure du flot sanguin en provenance de l'oreillette gauche ou de l'oreillette droite. Le système fonctionne ici comme une double pompe lors de la systole avec un circuit pulmonaire à basse pression et un circuit systémique à haute pression.
2.2.1. Généralités
Le travail cardiaque est le résultat d'un cycle contraction-relâchement du myocarde. Ce
cycle détermine: 1) une phase dite systolique, correspondant à la contraction et
engendrant une diminution du volume des chambres et une éjection du sang du ou des
ventricules; 2) une phase diastolique de relaxation musculaire correspondant à un retour
du volume des chambres aux conditions de repos et à leur remplissage. Ce cycle s'effectue
selon une certaine fréquence (pulsations/min). Le travail effectué détermine un volume
d'éjection (Qc) exprimé en ml ou en l et un débit cardiaque (
![]() ) généralement
exprimé en ml/min ou en l/min. Le débit cardiaque
) généralement
exprimé en ml/min ou en l/min. Le débit cardiaque
![]() est évidemment le produit de
la fréquence cardiaque par le volume d'éjection systolique.
est évidemment le produit de
la fréquence cardiaque par le volume d'éjection systolique.
![]() peut être déterminé in vivo,
sans avoir connaissance du volume d'éjection qu'il est souvent difficile de mesurer directement.
Dans ce cadre, on utilise le principe de Fick suivant lequel tout l'oxygène consommé par
un organisme est contenu dans le sang éjecté par le cœur. On peut donc connaître
le débit cardiaque en connaissant la consommation d'oxygène par minute (
peut être déterminé in vivo,
sans avoir connaissance du volume d'éjection qu'il est souvent difficile de mesurer directement.
Dans ce cadre, on utilise le principe de Fick suivant lequel tout l'oxygène consommé par
un organisme est contenu dans le sang éjecté par le cœur. On peut donc connaître
le débit cardiaque en connaissant la consommation d'oxygène par minute (
![]() ) et les
concentrations en O2 du sang artériel et veineux (CaO2 et CvO2):
) et les
concentrations en O2 du sang artériel et veineux (CaO2 et CvO2):

CaO2 peut être mesuré au niveau de n'importe quelle artère, elles ont en effet toutes la même pression artérielle en O2. La pression partielle en O2 du sang veineux peut largement dépendre de l'endroit choisi, en fonction du niveau de perfusion des organes se trouvant en amont. CvO2 sera donc pris le plus haut possible, au niveau de la veine cave ou du ventricule droit.
Le sang est éjecté sous une certaine pression résultant à la fois de la puissance et de la fréquence de contraction. La pression va varier entre un point haut systolique et un point bas diastolique.
Certains de ces paramètres sont repris pour différentes espèces dans le tableau 2-1.
Le travail cardiaque est essentiellement aérobie et nécessite une grande quantité d'énergie. Chez l'homme, on estime la consommation d'oxygène du cœur à 10% du QO2 total alors que le poids de cet organe n'excède pas 0.5% du poids total.
Le travail cardiaque par battement peut être estimé selon la formule empirique suivante:
![]()
avec
Qc: le volume d'éjection en ml
R: la pression artérielle moyenne en Kpa
m: la masse de sang déplacée exprimée en g (~Qc en ml)
V: vitesse moyenne du sang dans l'artère efférente en m/s
g: la constante d'accélération en ms-2
Ainsi, chez l'homme au repos, le travail du cœur gauche peut être estimé à:
![]()
En conditions d'activité importante, le travail augmente considérablement chez l'homme compte tenu d'une augmentation de la pression, du volume d'éjection et de la vitesse de déplacement du sang:

2.2.2. Commande et activité électrique - bruits du
cœur
La contraction du myocarde est commandée par des cellules produisant une excitation
électrique rythmique de façon totalement autonome (cfr. contraction
musculaire). Ces cellules constituent un tissu nodal ou rythmeur. Elles peuvent
être d'origine nerveuse et localisées en dehors du tissu contractile (rythmeurs
neurogéniques). C'est le cas chez différents invertébrés comme les crustacés
décapodes par exemple au niveau desquels un ganglion cardiaque d'une dizaine de cellules
joue le rôle de rythmeur. Lorsqu'il est séparé du cœur, celui-ci cesse de se
contracter alors que le ganglion isolé maintien une activité électrique spontanée.
Chez tous les vertébrés les rythmeurs sont dits myogéniques. Ils sont constitués de
cellules musculaires modifiées et font partie intégrante du tissu myocardique. Les
rythmeurs de mollusques et de tuniciers sont également myogéniques. On trouve des
rythmeurs soit myogéniques soit neurogéniques chez les annélides et les arthropodes.
L'activité électrique générée par les rythmeurs se propage sur le myocarde, déterminant la contraction. Chez les poissons, elle démarre au niveau du sinus veineux, en avant de l'atrium, les deux chambres étant pourvues d'un arrangement lâche de cellules myocardiques. Chez les mammifères, l'excitation naît au niveau d'un nœud dit sinusal ou sino-atrial situé sur le haut de l'oreillette droite. De là, elle s'étend sur les deux oreillettes et gagne le nœud atrioventriculaire. Elle est alors transmise par le faisceau de His au réseau de Purkinje, localisé au bas des ventricules. De là, l'excitation gagne les ventricules, de l'intérieur vers l'extérieur, du bas vers le haut (figure 2-8, A).
La variation d'activité électrique peut être suivie à l'aide d'électrodes externes. Ces variations, enregistrées sur des électrocardiogrammes, correspondent à des phases du cycle systole-diastole. Les tracés électrocardiographiques sont plus ou moins complexes en fonction de la complexité du système cardiaque des espèces considérées et de son système d'excitation.
L'activité cardiaque va également générer des modifications de pressions à différents endroits: oreillettes, ventricules, artères. Ces variations de pression vont par ailleurs déterminer des mouvements d'ouverture ou de fermeture de valves directionnelles se trouvant entre les différents compartiments du cœur. Si le cœur est suffisamment volumineux, les bruits résultant de ces mouvements, de même que certains bruits de passage du sang, peuvent être entendus par auscultation (figure 2-8, B) ou peuvent être enregistrés sous forme de phonogrammes.
Figure 2-8: Organisation du cœur des mammifères. A: Propagation de l'excitation. B: Valves mitrales et semi-lunaires: bruits du cœur. A: contraction des oreillettes, le sang passe dans les ventricules. B: contraction des ventricules, les valves mitrales se ferment occasionnant le 1er bruit du cœur. La pression monte dans le ventricule (C) provoquant l'ouverture des valves semi-lunaires (D) et le passage du sang dans l'artère aorte et l'artère pulmonaire. E: la pression baisse dans le ventricule, devient inférieure à celle existant dans les artères, ce qui provoque la fermeture des valves semi-lunaires, occasionnant le 2ème bruit du cœur. F: période de relaxation et de remplissage des oreillettes. G: le cycle recommence. D'après Florey 1968, modifié. (Voir aussi figure 2-12).
L'activité cardiaque peut donc être suivie par des mesures de fréquence de contraction, des tracés électrocardiographiques, des phonogrammes ou encore des mesures de pression et de débit à différents endroits. Récemment, on a ajouté à ces techniques, des techniques d'échographie et d'imagerie par résonance magnétique nucléaire. Ces technologies sont actuellement du domaine médical et n'ont pas encore été utilisée dans le domaine comparé, nous n'en discuterons donc pas ici.
La figure 2-9 reprend à titre d'exemple les électrocardiogrammes obtenus pour quelques espèces. En général, une onde marquée de dépolarisation correspond à la contraction de chaque chambre. On obtient ainsi un tracé simple avec un seul pic pour un criquet ou un crabe qui ont un cœur vésiculaire ou en forme de sac, à une seule chambre. Chez un bivalve, l'enregistrement montre deux ondes longues correspondant à la contraction de l'atrium d'abord, du ventricule ensuite. Chez les vertébrés, le tracé est plus complexe, formant un ensemble généralement noté P, QRS, T d'après les conventions adoptées pour les mammifères. P correspond à la dépolarisation associée à la contraction auriculaire. QRS est associé à la systole ventriculaire et la phase descendante de l’onde T correspond à la repolarisation ventriculaire (figure 2-10).
Figure 2-9: Exemples d'électrocardiogrammes.
Figure 2-10: Evolution de l'excitation sur un cœur de mammifère et phase correspondante du tracé électrocardiographique.
Ces phases de contraction induisent des variations de pression telles que celles reprises à titre d'exemple dans la figure 2-11 pour une truite et un requin. La figure 2-12 reprend par ailleurs l'évolution de différents paramètres au cours d'un cycle cardiaque au niveau de la partie gauche d'un cœur de mammifère. Comme le montre clairement cette figure, le premier bruit du cœur correspond à la fermeture de la valve mitrale (bicuspide), lors de la montée en pression du ventricule. Lorsque la pression intraventriculaire dépasse la pression dans l'aorte, la valve aortique (semi-lunaire) s'ouvre, déterminant le début de l'éjection du sang et la diminution du volume ventriculaire. La pression baisse alors dans le ventricule pour devenir inférieure à la pression aortique. Le volume systolique est minimum. La pression ventriculaire chute alors rapidement pour devenir inférieur à celle de l'oreillette, ce qui provoque l'ouverture de la valve mitrale et le remplissage du ventricule.
Figure 2-11: Evolution des pressions au niveau du cœur d'un requin et d'une truite en fonction du tracé électrocardiographique.
Figure 2-12: Evolution de différents paramètres lors du cycle cardiaque chez un mammifère.
2.2.3. Relation entre activité cardiaque, masse
corporelle et demande d'oxygène
La masse d'un organisme intervient de façon importante dans ses besoins énergétiques et
donc dans sa demande en oxygène (cfr. chapitre 3).
L'activité cardiaque étant liée à l'oxygénation des masses tissulaires, on pourrait
s'attendre à trouver une relation entre la masse des organismes et leur activité
cardiaque. Les conditions d'irrigation ainsi que les activités tissulaires sont cependant
très différentes en fonction des types d'organismes considérés. Il est dès lors
difficile d'extraire des traits généraux d'une comparaison activité cardiaque -
morphologie générale chez des espèces très éloignées les unes des autres. Cependant,
en considérant un groupe homogène, certaines constatations peuvent être faites.
Il existe par exemple une relation de proportionnalité directe entre la masse cardiaque et la masse corporelle au niveau d'espèces d'un même groupe zoologique. La figure 2-13 illustre ce point dans le cas des mammifères. Chez ceux-ci la pente de la droite est très voisine de 1 (0,98) et la masse cardiaque correspond à environ 0,59 % du poids de l'animal. Les données disponibles indiquent que chez les amphibiens et les reptiles, le poids du cœur correspond également à quelque 0,5 % du poids du corps. Le poids relatif du cœur est nettement inférieur chez les poissons ( 0,2 %) et significativement supérieur chez les oiseaux (0,8 %). La masse supérieure du cœur chez ces derniers pourrait être mise en relation avec le travail important qu'il doit effectuer lors du vol. A une masse plus importante correspond en effet un volume ventriculaire plus important et un volume d'éjection plus important.
| Figure 2-13: Relation entre la masse cardiaque et la masse corporelle chez les mammifères, y compris une série de primates. |
Il existe également, chez les vertébrés en tout cas, une relation entre la masse des organismes et la fréquence cardiaque. La fréquence est d'autant plus élevée que la masse est petite. Ce point est illustré dans la figure 2-14 pour les mammifères et les oiseaux. Par exemple, la fréquence au repos est de 25 pulsations/min chez un éléphant de 3000 kg alors qu'elle atteint 600 pulsations/min chez une musaraigne de 3 g. La relation masse-fréquence est la même que celle obtenue en étudiant le rapport consommation d'oxygène - masse (voir chapitre 3). Ceci indique que l'augmentation de fréquence cardiaque chez les animaux de petite taille est en relation directe avec leur consommation d'oxygène spécifique supérieure. Un point intéressant à remarquer ici et qui reste jusqu'à présent inexpliqué, est que l'augmentation de fréquence avec la masse des oiseaux est différente selon qu'ils sont au repos ou en vol. Il est possible que le travail de sustentation soit plus important pour un grand oiseau que pour un petit.
Figure 2-14: Relation entre la fréquence cardiaque et le poids des organismes. A: Chez les mammifères. D'après Schmidt-Nielsen 1983, modifié. B: Chez les oiseaux.
La demande en oxygène est aussi variable en fonction notamment, de l'activité. Dans ce cadre, tous les organismes répondent à la demande énergétique accrue en phase d'activité par une augmentation de fréquence cardiaque. Celle-ci peut s'accompagner d'une augmentation du volume d'éjection. Ce n'est pas toujours le cas comme nous le verrons par la suite (cfr. 4.1.2, ci-après).