

5.1.1. Le plasma sanguin
5.1.2. Les ťlťments figurťs
1) Les globules rouges (ťrythrocytes)
2) Les globules blancs (leucocytes)
3) Les thrombocytes
Comme nous l'avons signalť au chapitre 1, le sang se diffťrencie trŤs tŰt dans l'ťvolution animale comme un nouveau compartiment extracellulaire essentiel. Sa fonction initiale est de faciliter les ťchanges de matiŤre entre des masses tissulaires devenant trop importantes pour que ces ťchanges restent assurťs par diffusion. D'autres fonctions sont venues s'ajouter ŗ celle-ci, qui font du sang un "organe" essentiel ŗ plus d'un titre. Le sang va ainsi intervenir dans:
1) Les ťchanges gazeux (O2 et CO2) chez la plupart des
espŤces (voir chapitres 3 et 4).
2) Le transport d'ťlťments nutritifs et de produits terminaux du mťtabolisme
considťrťs comme "dťchets" (voir chapitres 5
et 6).
3) La communication entre cellules en acheminant les messagers chimiques, hormonaux et
autres, synthťtiser par diffťrents tissus (voir chapitre
9).
4) L'ťquilibre acide-base de l'organisme et le contrŰle du pH (voir chapitre
4).
5) Le mouvement au niveau de certains organes et la locomotion chez certaines espŤces par
modification de pression hydrostatique (voir chapitre
7).
6) Le transport de chaleur et la thermorťgulation au niveau de l'organisme chez les
homťothermes (oiseaux, mammifŤres) et au niveau de certains tissus chez diffťrents
pœcilothermes (autres espŤces) (voir chapitre
12).
7) La dťfense de l'organisme au niveau cellulaire contre des ťlťments
ťtrangers (voir chapitre 10).
8) L'osmorťgulation chez de nombreuses espŤces (voir chapitre
1).
9) La rťparation de dťchirures vasculaires par les phťnomŤnes d'hťmostase et de
coagulation (voir 5.2 ci-aprŤs).
Ainsi, le "liquide extratissulaire" (cfr. chapitre 1) des animaux est devenu au cours de l'ťvolution un support indispensable ŗ leur unitť fonctionnelle. Il est devenu le vťritable milieu de vie des cellules auxquelles il assure, chez les espŤces les plus ťvoluťes, une stabilitť d'environnement physico-chimique. Dans ce cadre, il apparaÓt comme un ťlťment majeur de la libťration des organismes vis ŗ vis des contraintes de l'environnement.
Le rŰle du sang dans l'osmorťgulation (8 dans la liste ci-dessus) a dťjŗ ťtť envisagť au chapitre 1. L'intervention du sang dans certaines des autres fonctions mentionnťes sera considťrťe ŗ l'occasion des chapitres spťcifiquement consacrťs ŗ ces fonctions. En ce qui concerne les mťcanismes de dťfense (point 7), il existe ŗ l'heure actuelle peu de donnťes comparatives permettant une intťgration dans un ensemble cohťrent. Chez les invertťbrťs, on a dťcrit de nombreux types de cellules sanguines qui phagocytent les dťbris cellulaires et tissulaires ainsi que les micro-organismes (voir aussi excrťtion). Certaines de ces cellules peuvent aussi sťcrťter une enveloppe kystique autour de corps ťtrangers, parasites notamment. Leur capacitť ŗ produire des protťines, les anticorps, capables de se fixer sur diffťrentes molťcules ťtrangŤres et d'induire contre celles-ci une phagocytose spťcifique (immunitaire) reste trŤs discutťe. Chez les vertťbrťs, par contre, ces rťactions dites immunitaires commencent ŗ Ítre connues de faÁon dťtaillťe. Elles seront briŤvement abordťes au chapitre 10. Par ailleurs, elles font maintenant classiquement l'objet de cours et d'ťcrits spťcifiques auxquels nous renvoyons le lecteur intťressť.
Nous n'envisagerons donc ici de faÁon trŤs brŤve que quelques notions concernant la composition globale du sang et son intervention dans la rťparation de dťchirures vasculaires (point 9).
Le sang est un liquide dans lequel peuvent flotter des cellules (ťlťments figurťs) en plus ou moins grand nombre. Le liquide une fois dťbarrassť des ťlťments figurťs porte gťnťralement le nom de plasma sanguin. Chez bon nombre d'invertťbrťs, la quantitť de cellules sanguines est trŤs faible, le sang se rťsume essentiellement au plasma. Par son origine, le sang doit Ítre considťrť comme un tissu mťsenchymateux dont les ťlťments cellulaires sont sťparťs par un liquide interstitiel, le plasma.
Le volume sanguin est extrÍmement variable (cfr. chapitre 1); il est beaucoup plus important chez la plupart des invertťbrťs que chez les vertťbrťs.
5.1.1. Le plasma sanguin
Le plasma est un liquide contenant en solution ou en suspension divers sels minťraux
(NaCl essentiellement, cfr. chapitre 1), ainsi que des
composťs organiques (substances nutritives, urťe, protťines). La nature et la
concentration des protťines plasmatiques sont trŤs variable d'un groupe zoologique ŗ
l'autre. Les donnťes concernant les invertťbrťs sont par ailleurs ťparses et peu
nombreuses. Nous nous concentrerons donc sur les vertťbrťs et plus particuliŤrement les
mammifŤres.
Chez les vertťbrťs, les protťines plasmatiques sont assez concentrťes et responsables du dťveloppement d'une pression oncotique non nťgligeable. Ce facteur joue un rŰle important dans les mouvements d'eau au niveau des capillaires (cfr. 3.3.2. ci avant). Ces protťines vont ťgalement intervenir dans le transport de nombreux ťlťments peu ou pas solubles. Elles vont ťgalement jouer un rŰle primordial dans les phťnomŤnes d'hťmostase et de coagulation (cfr. 5.1.2. ci-aprŤs) ainsi que dans les processus d'agrťgation des globules rouges qui vont influer sur la viscositť et donc les caractťristiques d'ťcoulement du sang.
Chez l'homme, la concentration en protťines atteint 70 ŗ 80 g/l. L'albumine reprťsente 60 % du total, une bonne part du reste, soit prŤs de 40 %, ťtant reprťsentť par des globulines (a1: 4 % - a2: 8 % - b: 12 % et g: 16 %) et diffťrents facteurs intervenant dans la coagulation du sang.
L'albumine va intervenir dans le transport de diffťrentes substances: bilirubine (voir plus loin) ou hormones par exemple. Elle peut ainsi servir de rťserve protťique en cas de dťficit nutritionnel.
Les a1, a2 et b globulines servent notamment au transport des lipides et de diverses vitamines (voir plus loin), de l'hťmoglobine (heptaglobine), du fer (transferrine), du cortisol (transcortine) et des cobalamines (transcobalamine).
Les immunoglobulines (Ig) font partie essentiellement des g - globulines. Elles constituent les substances immunitaires du plasma (anticorps). La plus importante d'entre elles en terme de concentration (7 ŗ 15 g/l) est l'IgB. Elle intervient avec les IgA, IgM, IgD et IgE dans les processus de dťfense de l'organisme contre l'intrusion de corps ťtrangers (cfr. immunologie).
5.1.2. Les ťlťments figurťs
Chez la plupart des organismes, on trouve des cellules capables de mouvements amiboÔdes
dont le rŰle essentiel est de phagocyter les dťbris cellulaires et les corps ťtrangers
(voir chapitre 6). Chez diffťrents invertťbrťs, les
ťlťments figurťs du sang se limitent ŗ ce type de cellules, en faible quantitť. Chez
les vertťbrťs, ces cellules entrent dans la catťgorie des leucocytes. On trouve par
ailleurs des thrombocytes et des ťrythrocytes (figure 2-31). Les
ťrythrocytes ont ťgalement ťtť appelťs globules rouges. Ils doivent leur couleur
rouge au pigment respiratoire colorť qu'ils contiennent, l'hťmoglobine (voir
chapitre 4). Les leucocytes, cellules incolores, sont
ťgalement appelťs globules blancs (figure 2-32), par opposition aux
globules rouges.
L'ensemble des ťlťments figurťs peut reprťsenter quelque 40 % du volume sanguin total chez les mammifŤres; ce qui correspond chez l'homme par exemple ŗ environ 5x106 particules par mm3. Le sang apparaÓt donc comme une suspension trŤs concentrťe dont les globules rouges sont les ťlťments majoritaires. Les ťrythrocytes vont donc intervenir de faÁon dťterminante dans les propriťtťs du sang (viscositť et dynamique d'ťcoulement dont elle dťpend). Dans certains cas, comme la leucťmie, le nombre de globules blancs peut toutefois s'accroÓtre suffisamment pour augmenter la viscositť de faÁon significative.
1) Les globules rouges (ťrythrocytes)
Le rŰle essentiel de ces cellules sanguines est le transport d'oxygŤne des surfaces
respiratoires vers les tissus (voir chapitre 4). On trouve
ces cellules chez quelques invertťbrťs et chez tous les vertťbrťs. Ils sont
exceptionnellement absents chez les Chaenichthyidťs, une famille de poissons
tťlťostťens antarctiques dťpourvus d'hťmoglobine.
Les globules rouges se prťsentent comme des cellules incapables de mouvements propres mais extrÍmement dťformables, dont la forme d'ťquilibre est un disque aplati, ovalisť (circulaire chez les cyclostomes et les mammifŤres). Comme nous l'avons vu, la dťformabilitť est, avec la capacitť d'empilement, une caractťristique importante intervenant dans la dynamique de l'ťcoulement sanguin (voir 3.2.). En conditions normales, la taille et le nombre de globules rouges sont pratiquement constants dans une mÍme espŤce mais varient beaucoup d'une espŤce ŗ l'autre et d'une classe de vertťbrťs ŗ l'autre. La taille ne varie en gťnťral pas au sein d'une mÍme espŤce, sauf cas pathologique. Le nombre par contre peut varier assez largement en fonction de diffťrents paramŤtres extťrieurs comme la disponibilitť de l'oxygŤne (voir chapitre 4).
Les globules rouges les plus petits et les plus nombreux se rencontrent chez les mammifŤres: homme: 5.106/mm3, diamŤtre 7,7 Ķ; chŤvre: 18.10 6/mm3, diamŤtre 4 Ķ. Ils sont nettement moins nombreux et plus grands chez les pœcilothermes. Ainsi, chez la grenouille, on en compte 6.105/mm3 de diamŤtre moyen 16 ŗ 22 Ķ. Au cours de leur diffťrenciation ŗ partir des cellules souches qui assurent leur formation et leur remplacement, les globules rouges de mammifŤres perdent leur chondriome, leur appareil de Golgi, leur centrosome et leur ARN. Le noyau est conservť dans tous les autres groupes; les mammifŤres sont les seuls organismes ŗ possťder des ťrythrocytes anuclťťs, appelťs aussi hťmaties.
Figure 2-31: Les ťlťments figurťs du sang de mammifŤres. Globules rouges: ťrythroblaste nuclťť (A) expulsant son noyau (B) pour donner naissance ŗ un ťrythrocyte anuclťť (hťmatie, C). M: mitochondrie. Vue en microscopie ťlectronique ŗ balayage d'hťmaties humaines.
2) Les globules blancs (leucocytes)
Alors que les globules rouges sont infťodťs au sang, les globules blancs sont des
cellules amiboÔdes qui empruntent la voie sanguine pour se rendre vers les endroits ou
ils sont nťcessaires. Leur nombre dans le sang est ťminemment variable, dťpendant des
facteurs capables de les mobiliser (digestion, infection, etc.). Il reste de toute faÁon
toujours largement infťrieur ŗ celui des globules rouges, le rapport rouge/blanc ťtant
de 5 ŗ 10 chez les poissons, 20 ŗ 70 chez les amphibiens, 30 ŗ 100 chez les reptiles,
70 ŗ 200 chez les oiseaux et 350 ŗ 2000 chez les mammifŤres. Cette augmentation de
rapport paraÓt plus en relation avec une augmentation du nombre de globules rouges
qu'avec une diminution du nombre de globules blancs. Il faut donc essentiellement y voir
une augmentation de l'efficacitť du systŤme de transport d'oxygŤne au cours de
l'ťvolution (cfr. chapitre 4).
Toujours nuclťťs et pourvus de tous les organites des cellules normales, les globules blancs se sťparent en deux groupes:
a) Les mononuclťaires qui sont de forme relativement sphťrique avec un noyau arrondi, non lobť. Ils se subdivisent en deux sous-groupes: les lymphocytes de petit diamŤtre (6 ŗ 8 Ķ chez l'homme) et les monocytes de plus grand diamŤtre (9 ŗ 12 Ķ chez l'homme). Cette division est peu nette chez les cyclostomes. Les lymphocytes sont responsables des rťactions immunitaires spťcifiques de dťfense (cfr. immunologie). Les monocytes sont en fait les prťcurseurs circulants des macrophages: cellules phagocytaires intervenant dans l'ťpuration du sang et l'ťlimination de cellules mortes et de dťbris de cellules (voir chapitre 6).
b) Les polynuclťaires qui ont un noyau lobť et polymorphe. Leur cytoplasme contient des granules spťcifiques qui leurs ont valu leur autre dťnomination de granulocytes. D'aprŤs la morphologie et la colorabilitť des granules on distingue des granulocytes eosinophiles, basophiles et hťtťrophiles (ou neutrophiles chez l'homme). Les proportions de ces trois catťgories varient d'une espŤce ŗ l'autre, les hťtťrophiles ťtant toutefois toujours de loin les plus nombreux (96 % chez l'homme).
Leur rŰle dans le sang est peu clair ŗ l'heure actuelle. Lorsqu'ils sont dans les tissus, ils paraissent intervenir largement dans la phagocytose des micro-organismes, les rťactions inflammatoires et l'immunitť antiparasitaire.
 |
 |
 |

 |
 |
 |
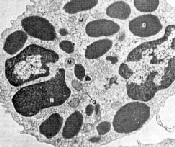
Figure 2-32: Les ťlťments figurťs du sang de mammifŤres. Globules blancs: leucocytes mononuclťaires (lymphocyte: A, monocyte: B) et polynuclťaires (granulocyte ťosinophile: D, basophile:E ou hťtťrophile: F). Plaquette ou thrombocyte: C. A/Ag: granules azurophiles, B: granules basophiles, C: centrosome, E: granules ťosinophiles, Ect: ectoplasme, G: Golgi, Gly: glycogŤne, Gr: granules, M: mitochondrie, N: noyau, Nu: nuclťole, Ps: pseudopode, R: ribosomes.
3) Les thrombocytes
Ils se prťsentent chez les non-mammaliens comme des cellules fusiformes nuclťťes,
souvent difficiles ŗ distinguer des petits lymphocytes. Chez les mammifŤres, ce sont des
structures anuclťťes, biconvexes, circulaires ou ovalaires, appelťes ťgalement
plaquettes.
Ces cellules interviennent dans le processus de coagulation du sang (voir ci-dessous).
Chez de nombreux invertťbrťs marins tels que certains vers ou holothuries, les pertes de sang suite ŗ blessure sont essentiellement contenues par des contractions du tťgument externe et, le cas ťchťant, des contractions des vaisseaux dťchirťs. Chez les autres invertťbrťs, les cellules sanguines ont la propriťtť de former des caillots en s'agglutinant. Chez certaines espŤces, crustacťs notamment, les cellules agglutinťes produisent un matťriel de type fibreux qui, s'amalgamant aux cellules, forme un coagulum. Ce matťriel paraÓt diffťrent de la fibrine intervenant dans la formation des caillots chez les vertťbrťs. Ces phťnomŤnes de coagulation sanguine restent peu ťtudiťs chez les invertťbrťs et il n'est guŤre possible ŗ l'heure actuelle de faire un examen comparatif sťrieux dťbouchant sur des donnťes gťnťrales.
Les mťcanismes intervenant ŗ la suite d'une dťchirure vasculaire ont surtout ťtť ťtudiťs chez les vertťbrťs et tout particuliŤrement chez les mammifŤres et l'homme. Chez ces espŤces, deux processus distincts dťmarrent en parallŤle: on assiste ainsi ŗ une hťmostase qui consiste en une obturation rapide de la dťchirure. Il y a par ailleurs une coagulation qui va aboutir ŗ la formation d'un thrombus fibreux bouchant complŤtement la dťchirure.
L'hťmostase met essentiellement en jeu les plaquettes (thrombocytes) se trouvant dans le sang circulant. Ces cellules vont s'agglutiner ŗ l'endroit de la lťsion au contact des fibres collagŤnes sous endothťliales mises ŗ nu par la dťchirure. Il y a ainsi formation d'un bouchon thrombocytaire (figures 2-33 et 2-34). Dans un phťnomŤne dit de mťtamorphose visqueuse, les thrombocytes agglutinťs vont alors se modifier sous l'influence d'un facteur sanguin, la thrombine (cfr. coagulation, ci-aprŤs), et libťrer, entre autres, de la sťrotonine qui induit une vasoconstriction locale. En outre, et dans le mÍme temps, la tunique interne du vaisseau lťsť s'enroule. Ces deux phťnomŤnes contribuent ŗ la poursuite de l'obturation vasculaire.
La coagulation intervient simultanťment ŗ l'hťmostase. Elle est dťclenchťe par la libťration de diffťrents facteurs par une voie exogŤne et une autre endogŤne (figure 2-33). Les diffťrentes ťtapes ne sont pas encore totalement dťfinies, briŤvement:
Dans la voie endogŤne, un facteur de coagulation plasmatique XII est activť par contact avec la zone lťsťe, vraisemblablement avec les fibres collagŤnes mises ŗ nu. Le facteur XIIa va lui-mÍme activer un facteur X.
Dans la voie exogŤne, les cellules endothťliales blessťes libŤrent des facteurs tissulaires qui, avec le facteur plasmatique VII et en prťsence notamment de Ca2+ et de vitamine K, vont ťgalement activer le facteur plasmatique X.
Figure 2-33: Quelques ťtapes importantes de l'hťmostase et de la coagulation du sang chez les mammifŤres. A.Ac: point d'action d'agents anticoagulants ou antithrombotiques.
Le facteur X activť, va, en prťsence de phospholipides des plaquettes, d'un facteur plasmatique V et de Ca2+, activer la prothrombine plasmatique (facteur II) en thrombine. La thrombine va: 1) activer le fibrinogŤne (facteur I) en fibrine; 2) initier la mťtamorphose visqueuse des thrombocytes; 3) activer un facteur plasmatique XIII de stabilisation de la fibrine. L'effet de la thrombine aboutit donc ŗ la formation de filaments de fibrine qui s'agglutinent en rťseau stable dans un bouchon thrombocytaire pour former le thrombus dťfinitif. C'est sur ce thrombus que va s'organiser la cicatrisation.
Dans le cas de lťsions minimes, c'est essentiellement la voie endogŤne qui intervient. Le systŤme exogŤne ne paraÓt Ítre mis en jeu que lors de lťsions plus importantes.
A l'heure actuelle, il n'est pas possible de rťpondre de faÁon complŤte aux questions de savoir comment la rťaction de coagulation ne s'ťtend pas ŗ tout le rťseau vasculaire et comment le thrombus se dissout aprŤs cicatrisation. En ce qui concerne ce dernier problŤme, diffťrents facteurs plasmatiques paraissent intervenir. Ainsi, le facteur plasmatique XII activť par les cellules endothťliales blessťes va, avec d'autres facteurs tissulaires et humoraux en rapport entre autres avec le dťficit en O2, activer le plasminogŤne sanguin en plasmine; ce qui va induire une fibrinolyse, c'est ŗ dire une lyse du caillot de fibrine.
Il existe par ailleurs dans le sang des substances antithrombiques (anticoagulantes). L'antithrombine 3 est la plus importante d'entre elles. Elle peut former des complexes avec la thrombine et le facteur X, inhibant ainsi leurs actions sur la formation de fibrine. La formation de ces complexes est renforcťe par l'hťparine, un agent anticoagulant bien connu extrait de diffťrents tissus animaux (foie, poumon, intestin, cerveau).
Les animaux se nourrissant de sang doivent sťcrťter des substances empÍchant la coagulation de celui-ci. Certains serpents disposent ťgalement dans leur venin de composťs capables d'interfťrer avec la coagulation sanguine (figure 2-33). Certains de ces produits inhibent le facteur X activť; c'est le cas de l'antistasine des sangsues et du facteur anticoagulant des tiques. D'autres bloquent l'action de la thrombine, empÍchant dŤs lors la polymťrisation de la fibrine ainsi que l'agrťgation des plaquettes: hirudine des sangsues et bothrojarasine dťcouverte rťcemment dans le venin d'un crotale d'Amťrique latine, Bothrops jararaca. Agiront ťgalement sur l'agrťgation plaquettaire mais ŗ d'autres niveaux la dťcorsine des sangsues et diverses substances des venins de serpents: thrombocytine, agrťgoserpentine ou bothrocitine par exemple. Le pouvoir anticoagulant de l'aspirine est ťgalement dŻ ŗ un effet sur l'agrťgation des plaquettes. D'autres substances encore agiront sur le fibrinogŤne, modifiant ses possibilitťs de polymťrisation; c'est le cas de l'hťmentine des sangsues ou de la crotalase. Certains agents enfin seront thrombolytiques, activant la formation de plasmine; c'est le cas du Bat-PA, un composť isolť ŗ partir des glandes salivaires d'une chauve-souris vampire sud amťricaine. La streptokinase, utilisťe actuellement pour dissoudre les caillots en cas d'infarctus du myocarde est ťgalement un thrombolytique efficace.



