 |
Figure 2-15: Empilement de globules rouges dans un vaisseau sanguin. |
3.3.1. Généralités
3.3.2. Echanges au niveau des capillaires
3.3.3. Les types de capillaires
Comme nous l'avons déjà signalé, le système circulatoire est ouvert chez la plupart des invertébrés alors qu'il est complètement fermé chez les vertébrés et chez quelques invertébrés considérés comme évolués (mollusques céphalopodes).
Dans le système fermé, le sang est amené à grande vitesse vers les tissus ou les échanges se font à travers un endothelium vasculaire à parois très minces, (vaisseaux capillaires). Le système vasculaire doit donc répondre ici à des exigences bien précises: par exemple, transport à haute pression et à grande vitesse exigeant résistance et élasticité pour certains tronçons, diamètre contrôlable permettant de diriger le flot sanguin sur d'autres tronçons, perméabilité élevée exigeant des parois minces sur d'autres encore. La circulation dans les vaisseaux sera par ailleurs régie par la mécanique des fluides.
Hagen et surtout Poiseuille ont fait au milieu du siècle dernier l'inventaire expérimental des facteurs qui influencent le débit d'un liquide newtonien en écoulement stationnaire à travers des tubes rigides de différentes sections. Un liquide newtonien est un liquide dont la viscosité ne varie pas avec le taux de frottement interne et reste constante quelle que soit la vitesse de l'écoulement laminaire. Comme nous le verrons, le sang est formé de particules et d'éléments cellulaires en suspension dans le plasma et sa viscosité est variable en fonction du débit, les vaisseaux ont par ailleurs un diamètre variable. Les lois de Poiseuille, Laplace et Bernouilli ne s'appliquent donc pas exactement au flot sanguin; nous ne nous étendrons dès lors pas longuement sur ces problèmes. Il est néanmoins intéressant de se rappeler quelques notions de base; à savoir: 1) le flot d'un liquide entre deux points est fonction de la différence de pression entre ces points et de la résistance à l'écoulement, 2) la résistance à l'écoulement est fonction de la viscosité du liquide, du diamètre de la tuyauterie et de sa longueur, 3) dans un système circulatoire, changer le diamètre engendre un changement de pression. Retenons par ailleurs avec Bayliss (Deformation and flow in biological systems - Frey - Wijssling editor, Amsterdam, 1952). "C'est une des ironies du progrès scientifique de constater que le débit de sang à travers un tube en verre, et, naturellement à travers les vaisseaux d'un animal, n'est pas proportionnel à la pression de perfusion sauf pour des débits élevés; il n'est pas non plus proportionnel à la puissance quatrième du rayon quand celui-ci est inférieur à 0,01 cm (comme c'est le cas dans la plupart des vaisseaux d'un animal vivant) et enfin il n'est pas toujours inversement proportionnel à la longueur du tube".
Le sang est un fluide non newtonien essentiellement parce que sa viscosité est variable en fonction de la vitesse de l'écoulement. Ce comportement, qualifié de rhéofluidifiant, est lié au fait que les globules rouges peuvent se déformer et s'agréger. Ces deux processus sont en relation étroite avec des propriétés du plasma et du globule rouge.
Le fibronogène, facteur I de la coagulation du sang (voir plus loin, section 5), est, avec les globulines du sang, responsable de la formation réversible de rouleaux de globules rouges (figure 2-15). Ces empilements sont surtout importants au niveau des gros troncs vasculaires et peuvent rendre compte de la grande augmentation de viscosité du sang observée à leur niveau. Ici, les caractéristiques de l'écoulement vont largement dépendre de la déformabilité des parois vasculaires. Au niveau des capillaires par contre, l'encombrement des globules rouges devient un élément essentiel (figure 2-16). On constate par ailleurs une baisse importante de la viscosité; les forces exercées par l'écoulement ont rompu les rouleaux. Ceci permet par ailleurs aux globules de s'aligner individuellement dans l'écoulement. Leur forme peut de plus se modifier considérablement, ce qui leurs permet de passer dans des capillaires de diamètre inférieur à leur diamètre moyen normal. Au niveau des veinules et des artérioles, un troisième phénomène intervient dans l'écoulement. Dans ces vaisseaux de diamètre relativement faible, on note une séparation de phase entre une région centrale riche en globules rouges et une couche liquide proche de la paroi. Cette couche exerce un effet lubrifiant facilitant l'écoulement des fluides dans la région centrale.
3.3.1. Généralités
Chez les espèces à système circulatoire fermé, on distingue des vaisseaux de
différents types. En partant du cœur, on rencontre tout d'abord les artères et les
artérioles. L'ensemble de ces vaisseaux, à l'exception de l'artère pulmonaire, amène
le sang oxygéné vers les tissus. Les capillaires, vaisseaux irriguant les tissus,
arrivent ensuite. Ils sont suivis des veinules et des veines, ramenant le sang
désoxygéné au cœur. Par ailleurs, l'artère pulmonaire amène le sang
désoxygéné du cœur vers les poumons et le sang oxygéné est ramené des poumons
vers le cœur par les veines pulmonaires. On distingue donc ainsi une "petite
circulation" pulmonaire d'une "grande circulation" ou "circulation
systémique" irriguant les tissus. Les caractéristiques des systèmes artériels,
veineux et capillaires de la grande circulation chez les mammifères sont résumées dans
les figures 2-17, 2-18 et dans les tableaux
2-2, 2-3.
Figure 2-17: A: Evolution de différents paramètres en fonction du type de vaisseau sanguin. B: Vue en microscopie électronique de deux artérioles (à droite) et de deux veines (à gauche). Remarquez l'épaisseur de la paroi des artérioles et l'importance de la couche de fibres musculaires lisses par rapport à celle des fibres élastiques, limitée à une fine lame du côté intérieur.
L'examen de ces données montre que :
1) Le système artériel comporte au départ du cœur des troncs de grand diamètre. Souples et résistants grâce à la présence de fibres élastiques, ils peuvent supporter les à-coups de pression dus au travail cardiaque et le passage d'un flot sanguin à haute pression. Les artérioles qui viennent ensuite sont caractérisées par la présence de couches de muscles lisses qui vont permettre des modifications de diamètre par variations de tonus. C'est à ce niveau que vont s'opérer les modifications de débit réglant l'importance de l'irrigation au niveau des différents organes.
2) Le système veineux est celui qui comporte le moins de vaisseaux mais c'est à ce niveau que se trouve le volume le plus important de sang (quelque 70 % du volume total, tableaux 2-2 et 2-3). C'est également à ce niveau que la pression est la plus basse. Elle va en diminuant régulièrement de l'artère aorte à la veine cave.
3) Le système capillaire constitue, avec les veinules, le système ayant la surface la plus importante et la perméabilité la plus élevée. C'est également à ce niveau que la vitesse du sang est la plus basse (de 0,1 à 0,3 cm/s contre 40 cm/s à la sortie du ventricule gauche) et que le nombre de vaisseaux est le plus élevé. C'est au niveau des capillaires que vont s'effectuer les échanges avec le liquide interstitiel environnant.
TABLEAU 2-2
Quelques caractéristiques du système vasculaire chez l'homme.

Figure 2-18: Caractéristiques des systèmes vasculaires. Evolution des surfaces et des vitesses d'écoulement en fonction du type de vaisseau chez un chien de 13 kg.
TABLEAU 2-3
Quelques caractéristiques du système vasculaire de la grande circulation chez le
chien.

3.3.2. Echanges au niveau des capillaires
Ces échanges font essentiellement intervenir un système dit de filtration-absorption
d'une part et un phénomène d'endocytose d'autre part. Il faut bien sur ajouter à ces
systèmes les systèmes de diffusion et de transport transmembranaire intervenant
classiquement dans les échanges de différentes espèces moléculaires au niveau de
toutes les membranes cellulaires Ces mécanismes occupent une place importante dans les
échanges ici comme ailleurs (cfr. physiologie cellulaire).
Le système d'endocytose sera considéré en détail dans le cadre du cours de physiologie cellulaire. Très brièvement, les molécules sont incorporées au niveau de la face apicale des cellules de l'endothelium dans des vésicules qui migrent vers la face basale au niveau de laquelle elles s'ouvrent, libérant leur contenu dans le liquide interstitiel.
Le système de filtration-absorption est schématisé dans la figure 2-19. Les forces qui interviennent dans ce type d'échanges sont d'une part la pression hydrostatique (filtration) générée par le travail cardiaque et qui s'exerce sur la paroi des capillaires et la pression osmotique (absorption) due aux molécules ne filtrant que mal au niveau des capillaires d'autre part. Il s'agit ici essentiellement des protéines plasmatiques. Chez les mammifères, leur concentration est de quelque 70 g/l de plasma et la pression osmotique qu'elles génèrent (dite pression oncotique) est d'environ 25 mmHg. 70 à 80 % de cette pression sont dus à l'albumine dont la concentration est élevée et le poids moléculaire relativement faible (70 000).
Figure 2-19: Echanges liquidiens entre plasma et liquide interstitiel. Le retour du liquide filtrant au système circulatoire peut également avoir lieu par le système lymphatique chez les espèces en possédant un. PH: pression hydrostatique. PO: pression oncotique.
La pression hydrostatique qui assure la filtration est évidemment la différence entre la pression hydrostatique à l'intérieur du capillaire et celle à l'extérieur de celui-ci, dans le liquide interstitiel. Cette pression a pu être estimée à 32 mmHg chez l'homme, au niveau d'une boucle capillaire d'un doigt. La pression de filtration à l'entrée d'un capillaire étant de 30 mmHg alors que la pression oncotique d'absorption n'est que de 25 mmHg, du liquide va filtrer du capillaire vers le milieu interstitiel entraînant avec lui les molécules dissoutes pour lesquelles la paroi de l'endothelium vasculaire est perméable (flot en masse, cfr. physiologie cellulaire). Le long du trajet du capillaire, la pression hydrostatique diminue alors que la pression oncotique ne varie guère. Au niveau de la portion veineuse de celui-ci, la pression hydrostatique devient inférieure à la pression oncotique. Il y a dès lors appel de liquide, suivant le gradient osmotique, du liquide interstitiel vers l'intérieur du capillaire.
Le système lymphatique peut, chez les espèces ou il est présent, participer également aux échanges en renvoyant vers le système circulatoire une partie du liquide filtrant des capillaires. Chez l'homme on estime ainsi que 20 litres de liquide sont filtrés en 24 heures par l'ensemble du réseau capillaire. Sur ces 20 litres, 18 litres sont réabsorbés directement au niveau des portions veineuses des capillaires et 2 litres retournent à la voie sanguine par le système lymphatique.
Les modifications de pression hydrostatique et/ou oncotique vont évidemment modifier l'équilibre des vitesses de filtration et d'absorption. Ce processus peut engendrer dans certains cas une augmentation significative de la quantité de liquide interstitiel avec pour résultat la formation d'œdèmes. Des œdèmes peuvent ainsi apparaître: 1) Lors d'une augmentation de la perméabilité aux protéines, par exemple occasionnée par l'histamine. Dans ces conditions, la pression oncotique diminue et la filtration domine. 2) Lors d'une diminution de la production de protéines sanguines, par exemple occasionnée par un mauvais fonctionnement hépatique. Dans ces conditions, la pression oncotique diminue et la filtration domine (œdème cirrhotique). 3) Lors d'une diminution du rappel du sang dans la portion veineuse des capillaires comme il en existe dans le cas de stase veineuse (insuffisance cardiaque ou manque d'activité des pompes veineuses sur les membres inférieurs). Dans ces conditions, la pression hydrostatique dans les capillaires augmente et dès lors la filtration domine.
L'organisation structurale des capillaires va évidemment influer grandement sur les vitesses d'échanges. Dans ce cadre, on distingue différents types de capillaires.
3.3.3. Les types de capillaires
On rencontre trois types de capillaires suivant les endroits irrigués et le
rôle fonctionnel qu'ils sont appelés à jouer (figure 2-20).
1) Les capillaires continus au niveau desquels la couche de cellules endothéliales est continue. Les cellules sont séparées par des espaces intercellulaires très minces ( 4 nm) offrant une certaine résistance à la filtration. La diffusion et l'endocytose interviennent ici de façon significative à côté du système de filtration-absorption. Ce type de capillaire forme l'essentiel du réseau.
2) Les capillaires fenestrés au niveau desquels les cellules présentent soit des amincissements importants du cytoplasme, mettant en contact les membranes plasmiques des faces apicales et basales (fenestrés fermés) soit des pores mettant en communication directe le milieu intravasculaire avec le liquide interstitiel (fenestrés ouverts). On trouve ces capillaires dans des endroits ou la filtration est importante tels que le glomérule rénal, le plexus choroïde, les villosités intestinales ou les glandes à sécrétion interne ou externe.
3) Les capillaires discontinus ou sinusoïdes au niveau desquels les espaces intercellulaires sont très larges, constituants de véritables brèches entre les cellules. Ces brèches permettent le passage de grosses molécules mais également de cellules sanguines (globules blancs et rouges). On trouve ce type de capillaires au niveau de la moelle osseuse, du foie et de la rate. Leur organisation explique que l'on puisse faire une transfusion à l'aide d'une aiguille implantée dans la moelle osseuse.
Le travail cardiaque détermine un flot sanguin qui va se distribuer dans le système circulatoire à une certaine pression et suivant un certain débit.
La pression étant directement fonction de l'activité cardiaque, on distinguera au niveau artériel une pression systolique et une pression diastolique correspondant respectivement aux phases de contraction et de relaxation du muscle ventriculaire. Par ailleurs, la pression, maximum à la sortie du cœur, au niveau des artères aorte et pulmonaire, va en diminuant pour chuter rapidement au niveau du réseau capillaire (figures 2-17, tableau 2-2). On va donc distinguer une circulation à haute pression comprenant essentiellement le système artériel allant du cœur aux capillaires irriguant les tissus d'une circulation à basse pression comprenant le système veineux ramenant le sang vers le cœur. Les pressions artérielles systoliques et diastoliques sont éminemment variables selon l'espèce envisagée (tableau 2-1). Dans certains groupes elles sont par ailleurs nettement plus élevées dans la grande circulation que dans la circulation respiratoire. C'est le cas au niveau de groupes évolués comme les mammifères ou les mollusques céphalopodes. Cette tendance évolutive se marque nettement au niveau des vertébrés comme le montrent les résultats de la figure 2-21. En fait comme le montre cette figure, c'est essentiellement la pression systémique qui devient plus élevée; ce qui permet un flot plus important et une ultrafiltration rénale plus efficace. Une pression basse au niveau respiratoire permet par ailleurs le maintien d'un épithélium très mince au niveau des surfaces d'échanges et donc une meilleure diffusion. Chez l'homme au repos par exemple, les pressions oscillent entre 120 et 80 mmHg dans la circulation systémique et entre 25 et 10 mmHg dans la circulation pulmonaire. Chez le poulpe les valeurs moyennes sont de 38 - 19 mmHg dans la grande circulation et de 7 - 3 dans la circulation branchiale. Chez ces espèces, la circulation basse pression comprend donc, outre le système veineux, l'ensemble du système de circulation respiratoire.
 |
Figure 2-21: Pression artérielle au niveau des circulations systémiques et respiratoires chez différents vertébrés. |
Le système à basse pression et particulièrement le système veineux sert de réservoir de sang grâce à sa grande capacité en rapport avec la grande déformabilité des veines (tableau 2-2). Le volume d'une veine est en effet extrêmement sensible à la pression transmurale. Les possibilités qu'ont les veines à parois fines de se collaber ou de gonfler jouent un rôle physiologique important en permettant des modifications considérables de volume sanguin en réponse à de très petits changements de la pression transmurale. Le système veineux peut ainsi stocker jusqu'à 80 % du volume sanguin total. Si par ailleurs le volume sanguin augmente, l'essentiel de l'augmentation ( 98 %) sera absorbée par le système veineux, 2 % seulement se répartissant dans le système artériel et capillaire. Inversement, une diminution de volémie se traduira d'abord presque exclusivement par une diminution de volume du système veineux. Ces phénomènes sont importants dans de nombreuses situations (pertes hémorragiques par exemple, voir plus loin). Par ailleurs la résistance que les veines opposent à l'écoulement est également modifiée par les changements de géométrie du profil veineux. Une veine collabée offre une résistance largement supérieure à une veine arrondie, largement ouverte. Ces phénomènes vont intervenir, entre autres, dans la régulation des effets gravitationnels sur la pression et l'écoulement du sang (voir plus loin).
Le flot sanguin se réparti au niveau des différents organes qui ne sont pas tous irrigués sous un même débit. Les données à ce sujet sont évidemment éminemment variables d'une espèce à l'autre. Généralement, les organes importants reçoivent une irrigation plus importante que les autres. Ainsi chez l'homme, une bonne part du débit cardiaque total (5,6 l/min) est répartie au niveau du rein, du foie, du cœur et du cerveau (3,6 l/min). Les 2 litres restant vont à la peau, au muscle et au reste des organes. Les différences de répartition sont mieux marquées encore lorsque les débits sont exprimés par unité de poids d'organe (tableau 2-4). Chez l'homme au repos, le rein reçoit quelque 4 l/min/kg et la peau 0,08 l/min/kg, soit 50 fois moins. Ces répartitions peuvent être modifiées et contrôlées par des phénomènes de vasodilatation et vasoconstriction pour faire face à différentes situations (effort musculaire, digestion, hémorragie etc., voir plus loin).
TABLEAU 2-4
Répartition du flot sanguin au niveau de différents organes chez l'homme
| Organe | Poids (kg) | Débit sanguin (l/min) | Débit sanguin (l/min/kg) |
| Rein | 0,3 | 1,2 | 4,0 |
| Foie | 1,5 | 1,4 | 0,9 |
| Cœur | 0,3 | 0,25 | 0,8 |
| Cerveau | 1,4 | 0,75 | 0,5 |
| Peau | 2,5 | 0,20 | 0,08 |
| Muscles | 29 | 0,90 | 0,03 |
| Reste | 35 | 0,90 | 0,03 |
Les pressions sanguines et les débits sont non seulement fonction de l'activité cardiaque ou des résistances périphériques mais encore de la position. Envisageons rapidement ce problème.
La pression existant au niveau d'une artère sera fonction de la pesanteur par suite du poids de la colonne de sang qui mène du cœur à l'organe considéré. Chez l'homme couché par exemple la pression artérielle au niveau du cœur, de la carotide et de l'artère pédieuse sera de 95 mmHg. Le passage à la position debout va diminuer la pression au niveau de la tête (carotide à 50 mmHg) et l'augmenter au niveau des pieds (artère pédieuse à 185 mmHg) (figure 2-22). Il faut noter que si la pression artérielle, au niveau de la tête par exemple, diminue de 45 mmHg, la pression veineuse de même que celle du liquide céphalo-rachidien diminue également d'autant; les échanges au niveau cérébral ne sont donc guère modifiés. Un sujet mis passivement de la position horizontale à la position verticale peut cependant présenter des vertiges transitoires. Ces vertiges ne sont donc pas à mettre en rapport avec les modifications des pressions artérielles et veineuses au niveau cérébral, mais plutôt avec la diminution du débit cardiaque que l'on observe dans ses conditions. Cette diminution n'est que transitoire et est rapidement régulée par vasoconstriction compensatrice (voir plus loin). Elle est due à la diminution du retour veineux qui suit le passage à la position verticale. Dans ce cadre, on notera par ailleurs qu'au niveau des pieds, l'augmentation de la pression transmurale peut générer une stase veineuse avec formation d'œdème si les jambes ne sont pas mises en mouvement. Au niveau des veines des membres inférieurs, comme des membres supérieurs d'ailleurs, l'endothelium vasculaire présente des replis qui jouent le rôle de valvules directionnelles. Au cours des mouvements, les contractions musculaires écrasent les veines, stimulant ainsi le retour veineux (pompe musculaire veineuse) et diminuant la pression. Chez l'homme debout au repos, la pression dans une veine du dos du pied est d'environ 90 mmHg. Cette pression tombe rapidement au tiers de cette valeur après quelques pas (figure 2-23).
Figure 2-22: Effets gravitationnels sur les pressions artérielles et veineuses chez l'homme au repos (pressions en mmHg).
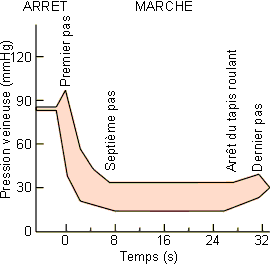 |
Figure 2-23: Effets de l'activation des pompes veineuses par la marche sur la pression dans la veine dorsale du pied chez l'homme. |
Les effets gravitationnels sont particulièrement marqués chez les grands animaux. Chez la girafe, la tête se trouve 2 m plus haut que le cœur. Pour maintenir à ce niveau une pression suffisante (100 mmHg), la pression artérielle au niveau cardiaque est très élevée (260 mmHg). Le système artériel des girafes possède des parois exceptionnellement épaisses capables de résister à ces pressions élevées. Pareillement les veines des membres inférieurs sont épaissies pour résister aux effets gravitationnels importants. Le système veineux est également équipé très largement de valvules directionnelles pour faciliter la circulation de retour. Ces valvules, au niveau des veines du cou, empêchent par ailleurs un afflux de sang et une stase veineuse au niveau cérébral lorsque la girafe se baisse pour boire, faisant passer sa tête de 2 m au-dessus du niveau du cœur à 2,5 m en dessous.
Les effets gravitationnels sur les pressions transmurales seront moins marqués chez les animaux aquatiques où ils sont compensés par des variations de pression hydrostatique. Une baleine passant de la position horizontale à la position verticale lors d'une plongée voit la pression transmurale au niveau des vaisseaux de sa tête augmenter. Cette augmentation est contrebalancée par une augmentation de pression hydrostatique, la hauteur de la colonne d'eau augmentant parallèlement à la hauteur de la colonne de sang.
Les effets posturaux ou gravitationnels sont en général très rapidement régulés et n'interviennent pas dans le contrôle normal du flot sanguin au niveau des différentes parties du corps. Ce contrôle est un élément essentiel de la physiologie de la circulation. Envisageons brièvement ses composantes majeures ainsi que quelques exemples dans lesquels il intervient de façon importante.








