4.2.1. Localisation - mode d'action
4.2.2. Caractťristiques et structures
4.2.3. Spťcificitť des effets
4.3.1. Gťnťralitťs introductives
4.3.2. Rťcepteurs couplťs ŗ des protťines G - SystŤme RCPG et seconds messagersa. les rťcepteurs
b. les protťines G
c. les systŤmes enzymatiques amplificateurs et les seconds messagers4.3.3. Rťcepteurs couplťs ŗ des tyrosines kinases cytoplasmiques - SystŤme RCTK
4.3.4. Rťcepteurs ŗ activitť tyrosine kinase - SystŤme RATK
4.3.5. Rťcepteurs ŗ activitť autre que tyrosine kinase
Les modulations
díactivitť induites par les sťmiomolťcules envisagťes dans ce chapitre
font intervenir un transfert díinformation du milieu extracellulaire vers le
milieu intracellulaire. La premiŤre ťtape de ce transfert fait toujours
intervenir une association avec un rťcepteur spťcifique. Aucune des sťmiomolťcules
considťrťes ici níagit en effet
directement sur les systŤmes effecteurs des cellules-cibles (enzymes,
transporteurs, canaux, gŤnes). Les rťcepteurs impliquťs sont membranaires
lorsque les molťcules passent difficilement la membrane plasmique (hormones et
parahormones peptidiques, cytokines, prostanoÔdes). Ils sont intracellulaires
pour les molťcules plus lipophiles et dont la permťabilitť membranaire est
plus ťlevťe (hormones stťroÔdes) ou encore pour les molťcules endocytťes
(hormones thyroÔdiennes iodťes). La distinction n'est toutefois pas toujours
aussi nette, on trouve en effet des rťcepteurs cytoplasmiques pour diffťrents
modulateurs dont les effets majeurs font intervenir des rťcepteurs
membranaires. De mÍme, diffťrentes hormones stťroÔdes ont, ŗ cŰtť de rťcepteurs
intracellulaires, des rťcepteurs membranaires. Le rŰle de ces rťcepteurs
"secondaires" reste en gťnťral peu clair jusqu'ŗ prťsent : ils ne
seront donc pas considťrťs plus en dťtail ici.
En gťnťral, les rťcepteurs intracellulaires activťs peuvent moduler directement líactivitť de leurs systŤmes-cibles. La modulation par rťcepteur membranaire doit par contre faire intervenir une cascade de signalisation permettant le transfert du message de la membrane au site intracellulaire du systŤme-cible. Envisageons maintenant rapidement ces diffťrents mťcanismes.
4.2.1. Localisation - mode
d'action
Les hormones stťroÔdes ou thyroÔ≠diennes
forment avec leurs rťcepteurs intracellulaires des complexes qui, d'une faÁon
encore mal connue, vont activer la synthŤse de diffťrentes protťines au
niveau gťnique. Ils sont essentiellement localisťs dans le noyau des
cellules-cibles et leur effet majeur paraÓt se situer au niveau díune rťgulation
de la transcription des gŤnes (figure 9-29). Il paraÓt cependant clair que d'autres
processus, comme par exemple une modulation de líactivitť translationnelle
pourraient ťgalement Ítre impliquťs. Des effets non gťniques peuvent par
ailleurs Ítre pris en considťration dans certains cas, tels que modification
de dťgradation ou de stockage de la protťine, modification d'activitť par
phosphorylation ou glycosylation.
Le schťma le plus communťment admis ŗ l'heure actuelle considŤre que l'essentiel des rťcepteurs inactifs se trouve au niveau du noyau, associť ŗ des protťines non-histoniques de la chromatine. L'association avec l'hormone libŤre le rťcepteur et le complexe HR actif peut se fixer ŗ un ťlťment de la rťgion rťgulatrice (5'-flanking) d'une unitť de transcription, participant ainsi, ťventuellement avec d'autres ťlťments modulateurs, ŗ son activation ou au contrŰle de sa vitesse de transcription (figure 9-29). Ce modŤle gťnťral parait valable ŗ la fois pour les hormones stťroÔdes et les hormones thyroÔdiennes. Une exception a toutefois ťtť clairement dťcrite ŗ ce jour ; il s'agit de l'ťlťment rťgulateur glucocorticostťroÔde du gŤne de l'hormone de croissance qui parait situť sur le premier intron de l'unitť structurale et non dans la rťgion rťgulatrice.
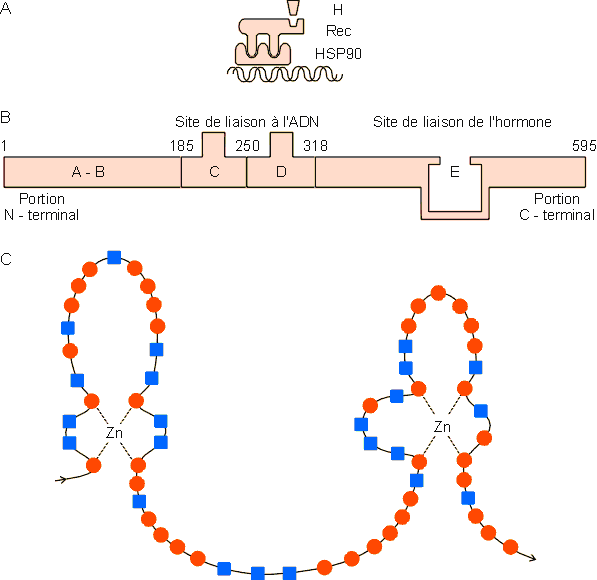
Figure 9-29 : Reprťsentation schťmatique du rŰle des rťcepteurs intracellulaires dans le mode d'action des hormones stťroÔdes. H : hormone, Rec : rťcepteur, HSP90 : protťine associťe au Rec bloquant son association ŗ l'ADN. Le complexe hormone-rťcepteur une fois libťrť du HSP90 peut se lier ŗ un ťlťment de rťponse hormonal (HRE) se trouvant sur la rťgion 5' rťgulatrice d'une unitť transcriptionnelle. Cette liaison dťterminera, avec l'activation ťventuelle d'autres ťlťments de rťponse (RE), la vitesse de transcription d'une sťrie de gŤnes de structure. P : promoteur TATA. Dťtails dans le texte.
4.2.2. Caractťristiques et
structures
Les rťcepteurs aux hormones stťroÔdes et thyroÔdiennes paraissent appartenir
ŗ une mÍme grande famille. On en a proposť un classement en 6 catťgories sur
base des spťcificitťs de leur liaison avec diffťrents ligands : hormones,
agonistes et antagonistes. Nombre d'entre eux ont maintenant ťtť identifiťs
chez les mammifŤres et clonťs ; l'ťtude de leur sťquence a permis de
les grouper en deux classes : les rťcepteurs de classe 1 comprenant les
rťcepteurs aux stťroÔdes des surrťnales (gluco et minťralocorticoÔdes)
ainsi que les rťcepteurs ŗ la progestťrone et ceux aux androgŤnes - les
rťcepteurs de classe 2 comprenant les rťcepteurs aux ústrogŤnes et ceux aux
hormones thyroÔdiennes.
Les rťcepteurs aux stťroÔdes ont ťtť les plus ťtudiťs. Ils ont des structures trŤs proches avec, notamment des sites de liaison ŗ l'ADN comportant deux complexes de coordination de type "doigt de zinc". Ils se prťsentent sous forme de gros hťtťro-oligomŤres comprenant des sous-unitťs protťiques associťes ŗ une unitť de type HSP 90 inhibant leur association au site de liaison de l'ADN. La liaison hormone-rťcepteur induit le relarguage de cette unitť (figure 9-30).
Figure 9-30 : A : Reprťsentation schťmatique d'un rťcepteur ŗ hormone stťroÔde. Ces rťcepteurs se prťsentent tous sous la forme d'hťtťro - oligomŤres paraissant associťs ŗ une protťine de type HSP90 (Heat Shock Protein - voir aussi chapitre 12) qui empÍche l'association du rťcepteur non activť avec l'ADN. La liaison de l'hormone au rťcepteur permet cette association en libťrant le HSP90. B : Structure linťarisťe du rťcepteur ŗ l'ústradiol humain, un ensemble de 595 acides aminťs avec des domaines centraux C et D contenant le site de liaison ŗ l'ADN et un domaine C-terminal E contenant le site de liaison du ligand. Ce site parait consister en un ensemble de 12 structures a hťlicoÔdales formant une cavitť au niveau de laquelle les interactions entre ústradiol, Glu 353, Arg 394 et His 524 jouent un rŰle essentiel dans la liaison (voir Brzozowski et al., Nature 387, 753, 1997). Ce type de structure parait gťnťral au niveau des rťcepteurs ŗ hormone stťroÔde. C : Les sites de liaison ŗ l'ADN paraissent ťgalement de structure trŤs proche au niveau des diffťrents rťcepteurs. Ils prťsentent deux complexes de coordination formant des doigts de zinc qui jouent vraisemblablement un rŰle important dans la liaison du rťcepteur activť ŗ son ťlťment de rťponse sur l'ADN. La partie C de la figure compare les sites de liaison ŗ l'ADN pour les rťcepteurs ŗ l'ústradiol, ŗ la progestťrone et au corticostťrol. Les acides aminťs communs aux diffťrents rťcepteurs sont reprťsentťs par l, les autres par n.
4.2.3. Spťcificitť des effets
Il ne paraÓt pas y avoir de mťcanisme de transport transmembranaire
spťcifique aux hormones stťroÔdes ou thyroÔdiennes. Par ailleurs, la
permťabilitť des cellules-cibles ŗ ces hormones ne paraÓt pas
particuliŤrement ťlevťe. Elles diffusent donc dans tous les types
cellulaires ; la capacitť de rťponse spťcifique des cellules-cibles
dťpend en fait de la prťsence de rťcepteurs dans les cellules plutŰt que de
l'accessibilitť de ces cellules ŗ l'hormone.
La spťcificitť d'effets en fonction du type de cellule-cible paraÓt, elle, liťe ŗ des diffťrences dans les unitťs de transcription activťes plutŰt qu'ŗ l'existence de diffťrents types de rťcepteurs activant chacun des unitťs donnťes. Ainsi par exemple, les rťcepteurs aux glucocorticoÔdes ou ŗ la testostťrone sont les mÍmes dans les diffťrents types de cellules-cibles. La similitude des rťcepteurs peut rendre compte du syndrome de fťminisation testiculaire observť chez de nombreux mammifŤres au niveau desquels aucun des caractŤres sexuels m‚les n'apparaÓt bien que les testicules soient prťsents et sťcrŤtent de la testostťrone en quantitť suffisante. L'anomalie rťside ici en un dťfaut de synthŤse du rťcepteur ; il en rťsulte des individus gťnťtiquement m‚les mais au niveau desquels l'expression phťnotypique de l'ensemble des traits masculins est bloquťe par manque du rťcepteur requis.
En gťnťral, l'activation d'un systŤme rťcepteur module la transcription de quelques dizaines ŗ quelques centaines de gŤnes, le plus souvent semble-t-il en un systŤme de cascade. De ce cadre, une activation "primaire" porte sur un petit nombre de gŤnes et induit la production de quelques protťines modulatrices qui vont elles-mÍmes activer, dans une rťponse "secondaire", d'autres unitťs transcriptionnelles. Ainsi par exemple, l'hormone de mue de la drosophile provoque, dans les 10 minutes suivant son injection, l'apparition de 6 nouveaux sites de synthŤse de RNA sur les chromosomes gťants. AprŤs dťlais, on constate l'apparition de plus de 100 autres sites ; l'ensemble aboutissant ŗ la formation de trŤs nombreuses architectures protťiques nouvelles.
Au niveau des cellules animales, l'organisation en opťrons prťsente chez les bactťries n'existe pas ; les sťquences codant pour diffťrentes unitťs d'une mÍme structure protťique peuvent mÍme se trouver sur des chromosomes diffťrents (hťmoglobine par exemple). Il doit donc exister un mťcanisme sťlectionnant les unitťs de transcription ŗ activer en fonction des effets ŗ obtenir. On ne connaÓt ŗ l'heure actuelle pratiquement rien des facteurs qui pourraient dťterminer la "compťtence" des gŤnes devant Ítre rťgulťs dans un cadre donnť. Il semble que le jeu de diffťrents facteurs de transcription intervenant non seulement au niveau des segments rťgulateurs mais aussi au niveau des introns doive Ítre pris en considťration. Les segments de DNA ainsi impliquťs dans ce cadre pourraient soit activer la transcription de certains gŤnes (''enhancer segments") soit encore la bloquer ("silencer segments").
4.3.1. Gťnťralitťs
introductives
La majeure partie des sťmiomolťcules envisagťes jusqu'ici font intervenir des
rťcepteurs membranaires. Ces rťcepteurs occupent donc une place essentielle
dans les systŤmes de signalisation intercellulaires. Ils prťsentent tous
certaines caractťristiques communes, notamment : un domaine extracellulaire
pouvant fixer spťcifiquement le modulateur et un domaine membranaire
intervenant dans la transduction du signal du milieu extracellulaire au milieu
intracellulaire. Dans la plupart des cas, on trouve ťgalement un domaine
intracellulaire assurant la transmission du signal ŗ un ou plusieurs systŤmes
effecteurs ; cette transmission fait gťnťralement intervenir une cascade
plus ou moins longue des systŤmes relais kinasiques ou phosphatasiques.
On regroupe habituellement les rťcepteurs membranaires en
cinq catťgories :
- les rťcepteurs dont l'organisation structurelle forme un
canal.
- les rťcepteurs couplťs ŗ des kinases de signalisation
intracellulaires par l'intermťdiaire de protťines dites G.
- les rťcepteurs couplťs directement ŗ des enzymes
cytoplasmiques de signalisation.
- les rťcepteurs possťdant une activitť enzymatique
propre, gťnťralement de type kinasique ou phosphatasique.
- les rťcepteurs du type anticorps du systŤme immunitaire.
Les rťcepteurs de type anticorps sont spťcifiques au systŤme immunitaire ; ils seront donc envisagťs au chapitre 10 traitant des mťcanismes de dťfense.
Les rťcepteurs "canaux" interviennent essentiellement dans la transmission de signaux relatifs ŗ la modulation du potentiel de membrane et ŗ sa propagation au niveau synaptique. Ils sont donc considťrťs au chapitre 8 traitant des relations nerveuses.
Les trois autres catťgories de rťcepteurs interviennent dans la transmission de nombreux messages hormonaux, parahormonaux ou relevant de modulateurs locaux qui nous intťressent ici. Envisageons-les donc rapidement de maniŤre successive.
4.3.2. Rťcepteurs couplťs ŗ
des protťines G - SystŤme RCPG et seconds messagers
Le systŤme RCPG est impliquť dans la transmission au niveau
intracellulaire des messages de trŤs nombreux modulateurs : hormones
peptidiques (mis ŗ part l'insuline, cfr. 4.3.4,
líhormone de croissance et la prolactine, cfr.4.3.3),
catťcholamines, prostaglandines, leucotriŤnes et autres eicosanoÔdes ainsi
que diffťrentes cytokines.
Il est probablement le systŤme le mieux connu ŗ l'heure actuelle, ayant servi dans les annťes 50 de premier modŤle d'ťtude concernant la transmission d'un signal hormonal. Il s'agissait en fait de líťtude des effets de la noradrťnaline sur la glycogťnolyse au niveau de cellules hťpatiques (cfr. 2.3.4 et 4.4Ö). Ces recherches ont permis ŗ Sutherland, leur auteur, de dťvelopper le concept de "second messager" qui lui valu le prix Nobel de Mťdecine en 1960. Dans ce systŤme, l'activation d'un rťcepteur membranaire par un "premier messager" (la noradrťnaline) induit dans le cytoplasme des cellules-cibles la production d'un "second messager" qui ŗ son tour va activer diffťrents processus intervenant dans l'effet de l'hormone. Le second messager intervenant dans le cas prťsent est l'AMP cyclique (AMPc). On en a depuis lors dťcouvert plusieurs autres ; leur production fait chaque fois intervenir une protťine membranaire dite de type G ; ainsi nommťe parce qu'un nuclťotide guanylique, le GTP, participe ŗ son activation (voir plus loin et figure 9-31).
Le systŤme RCPG concerne donc une sťquence de transduction d'un message vťhiculť dans le milieu extracellulaire par un modulateur "premier messager" vers le milieu intracellulaire et comprenant : 1) la liaison du ligand (modulateur) ŗ son rťcepteur spťcifique. 2) L'activation d'une protťine G. 3) L'activation d'un systŤme enzymatique effecteur impliquť dans la production du "second messager" (figure 9-31).
Le systŤme enzymatique effecteur des protťines G est parfois appelť amplificateur puisqu'en fait son activation ŗ partir d'un seul complexe ligand-rťcepteur aboutit ŗ la formation de nombreuses molťcules de second messager, amplifiant donc en quelque sorte le signal de dťpart. Dans certains cas, comme nous le verrons par la suite, la liaison ligand-rťcepteur aboutit ŗ une inhibition de l'activitť du systŤme amplificateur au lieu d'une stimulation. Il y a donc dans ces conditions une diminution de la quantitť de second messager au lieu d'une augmentation (figure 9-31).
Figure 9-31 : Le systŤme de signalisation RCPG et la formation des "seconds messagers". Dťtails dans le texte. R : rťcepteur spťcifique ŗ un premier messager ; PGs, PGi : protťine G stimulatrice ou inhibitrice (cette derniŤre dans certains cas seulement) ; SA : systŤme enzymatique amplificateur ; PK : Kinase de protťine ; Ca2+ : Calcium.
Les diffťrents second messagers produits vont induire des effets physiologiques par l'intermťdiaire d'une modulation 1) de l'activitť de diffťrentes kinases, enzymes activant ou inhibant par phosphorylation une variťtť de protťines prťsentant des activitťs biologiques (enzymes, canaux, transporteurs) ou encore 2) de la concentration en calcium intracellulaire, variation induisant ŗ son tour une modulation de l'activitť de diffťrentes protťines.
Envisageons maintenant de maniŤre un peu plus dťtaillťe les diffťrentes composantes du systŤme RCPG.
a. les rťcepteurs
Les rťcepteurs du systŤme RCPG constituent sans nul doute la famille la plus
nombreuse des rťcepteurs membranaires. Ils restent ŗ l'heure actuelle loin
d'Ítre tous complŤtement caractťrisťs mais les donnťes obtenues jusqu'ŗ
prťsent montrent qu'ils prťsentent tous des caractťristiques structurales
fondamentales communes ŗ cŰtť des spťcificitťs de configuration
nťcessaires notamment ŗ la liaison avec un modulateur donnť. Ils sont en
effet tous constituťs d'une seule chaÓne polypeptidique possťdant 7 segments
transmembranaires en hťlice a (figure
9-32). L'extrťmitť N-terminale est localisťe dans le milieu
extracellulaire et porte des sites de glycosylation. L'extrťmitť C-terminale
est cytosolique et prťsente des sites de phosphorylation. La boucle
cytoplasmique C3 parait Ítre un ťlťment clef dans l'interaction avec les
protťines G.
Contrairement ŗ ce que l'on aurait pu imaginer, de nombreux ligands de petite taille ne se fixent pas sur la partie N-terminale, aisťment accessible dans le milieu extracellulaire, mais plutŰt entre les hťlices transmembranaires. L'encombrement stťrique des ligands de grande taille empÍche ce type de fixation qui implique alors soit les boucles extramembranaires de la partie N-terminale, soit l'interface entre ces boucles et la membrane.
Le rŰle des rťcepteurs RCPG est en dťfinitive triple : 1) ils fixent spťcifiquement un modulateur donnť, 2) ils transmettent le signal au sein de la membrane, 3) ils interagissent avec des protťines G permettant ainsi la production de seconds messagers.
Figure 9-32 : Topologie proposťe pour les rťcepteurs de la famille RCPG.
b. les protťines G
Les protťines G interagissent avec les rťcepteurs RCPG au niveau de la face
cytoplasmique de la membrane. Il s'agit d'une interaction trŤs spťcifique, un
rťcepteur donnť activant une protťine G donnťe. Il y a donc de trŤs
nombreuses protťines G. Elles se prťsentent toutes comme des hťtťrotrimŤres
comportant 3 sous-unitťs : a , b
et g . On les distingue essentiellement par la nature
de leurs sous-unitťs a , beaucoup plus nombreuses et
variťes que les deux autres qui restent semblables dans de nombreux cas. La
sous-unitť a lie par ailleurs un nuclťotide
guanylique et prťsente une activitť GTPasique qui vont jouer un rŰle
essentiel dans l'activation et l'inactivation des protťines G (figure
9-33).
A l'ťtat inactif, la sous-unitť a lie en fait un GDP et prťsente une forte affinitť pour les sous-unitťs b et g ; la protťine G existe donc alors sous une forme a GDP b g (figure 9-33 Ā ). L'activation du rťcepteur permet la liaison de la sous-unitť a avec ce dernier ; elle induit ťgalement une modification d'affinitť pour le nuclťotide guanylique. Le GDP est ainsi remplacť par un GTP (figure 9-33 ā ). A l'inverse de a GDP, a GTP prťsente une affinitť faible pour le rťcepteur et pour les sous-unitťs b et g . Il va donc se dissocier de ses structures et pourra alors activer un systŤme amplificateur (figure 9-33 É ). a GTP restera actif tant que son activitť GTPasique n'aura pas hydrolysť le GTP en GDP, restituant donc la forme a GDP qui se rťassociera alors avec les sous-unitťs b et g et ensuite avec un nouveau rťcepteur activť pour recommencer un cycle (figure 9-33 Ą ).
Le rťcepteur restť actif lors du dťpart de a GDP peut de son cŰtť fixer un nouveau complexe inactif a GDP b g pour dťmarrer un autre cycle. On voit donc, dans ce contexte, que l'amplification du signal portť par un modulateur extracellulaire a lieu en fait ŗ deux niveaux ; tout d'abord au niveau du rťcepteur activť qui peut activer de nombreuses protťines G les unes ŗ la suite des autres ; ensuite au niveau de la sous-unitť a GTP qui peut maintenir le systŤme amplificateur activť tant que le GTP n'est pas hydrolysť en GDP (figure 9-33).
Figure 9-33 : Cycle d'activation et inactivation des protťines G dans la transmission du signal d'un rťcepteur RCPG ŗ un systŤme amplificateur. L : ligand (modulateur) ; R : rťcepteur ; a, b, g : sous-unitťs d'une protťine G. Dťtails dans le texte.
c. les systŤmes enzymatiques
amplificateurs et les seconds messagers
L'activation des protťines G conduit ŗ la modulation de l'activitť de
systŤmes effecteurs amplificateurs (enzymes ou canaux) aboutissant ŗ une
modification de concentration de messagers intracellulaires (seconds messagers).
Avant d'envisager plus en dťtail ce systŤme, rappelons que les mťcanismes
intervenant dans le contrŰle de canaux ioniques en rapport avec la gťnťration
de potentiels d'action ou de jonction sont envisagťs au chapitre 8 traitant du systŤme nerveux.
Les messagers les plus connus ŗ l'heure actuelle sont soit des nuclťotides cycliques (AMPc et GMPc) soit des dťrivťs de certains lipides membranaires : diacylglycťrol (DAG) et inositol triphosphate (IP3), dťrivťs du phosphoinositol biphosphate - lysophospholipides dťrivťs de la phosphatidylcholine. Les enzymes impliquťs dans leur formation sont respectivement des adťnylyl-cyclases pour l'AMPc, des guanylyl-cyclases pour le GMPc, des phospholipases C pour l'IP3 et le DAG et des phospholipases A2 pour les lysophospholipides (figure 9-34). L'activation de phospholipases D peut ťgalement produire du DAG ŗ partir de phosphatidylcholine membranaire (voir plus loin).
Les cibles intracellulaires directes de ces seconds messagers ne sont pas encore toutes clairement dťfinies. Signalons simplement ici les protťines kinases A, G et C, sensibles respectivement ŗ l'AMPc, au GMPc et au diacylglycťrol. L'IP3 agira lui en induisant une mobilisation du Ca2+ ŗ partir des rťserves du rťticulum endoplasmique. Il active en fait l'ouverture de canaux ŗ Ca2+ au niveau de ces structures, favorisant ainsi sa libťration et son accumulation dans le cytoplasme. Le Ca2+ peut donc Ítre considťrť comme une sorte de "troisiŤme messager" ; il agira soit directement sur l'activitť de diffťrents systŤmes effecteurs soit via diffťrents types de protťines fixant le Ca2+ (voir plus loin et figure 9-34).
Figure 9-34 : Les systŤmes enzymatiques amplificateurs, les "seconds" et "troisiŤmes" messagers. Les phospholipases A2 sont activťes en prťsence de Ca2+ ; elles s'associent alors ŗ la membrane pour produire des lysophospholipides ŗ partir de phosphatidylcholine membranaire. DAG : diacylglycťrol ; G : protťine G ; IP3 : inositol triphosphate ; L : ligand (modulateur) ; PC : phosphatidylcholine ; Pi P2 : phosphatidylinositol-biphosphate ; PK : Kinases de protťines. Dťtails dans le texte.
La rťgulation de la concentration intracellulaire en second messager peut mettre en jeu non seulement l'activitť des systŤmes enzymatiques amplificateurs responsables de leur synthŤse mais encore l'activitť des systŤmes responsables de leur dťgradation. Dans ce cadre, la dťgradation des nuclťotides cycliques AMPc et GMPc fait intervenir des phosphodiestťrases qui les dťcyclisent en nuclťotides 5' correspondants (figure 9-35). On ne connaÓt ŗ l'heure actuelle que trŤs peu de choses concernant la rťgulation de ces voies mťtaboliques. Rappelons, par ailleurs, que, dans le cas particulier de l'adťnylyl-cyclase, il existe des protťines G (Gi) qui sont inhibitrices de l'activitť de l'enzyme et qui vont donc induire une diminution de la teneur en AMPc. Rappelons ťgalement que le DAG peut Ítre produit suite ŗ activation de phospholipases C ou D. Comme le montre la figure 9-36, cette production est nettement plus lente lors de l'activation de la phospholipase D. Les deux types de production du DAG interviennent donc vraisemblablement dans des types de rťgulations diffťrents.
Figure 9-35 : Principaux seconds messagers et voie de synthŤse.
Figure 9-36 : A : Evolution du diacylglycťrol (DAG) et de l'activitť de la PKC suite ŗ activation de phospholipases C (óóó, DAG produit ŗ partir de PIP2 ) ou de phospholipases D (óóó, DAG produit ŗ partir de PC). B : L'activation des phospholipases C induit ťgalement une augmentation rapide du Ca++ (via IP3 ) responsable d'une activation des phospholipases A2 et d'une augmentation lente des lysoPC.
Quelques exemples de rťgulation impliquant certains des seconds messagers considťrťs ci-dessus seront envisagťs dans la section suivante (4.4).
4.3.3. Rťcepteurs couplťs ŗ
des tyrosines kinases cytoplasmiques - SystŤme RCTK
Cette catťgorie de rťcepteurs intervient essentiellement dans la transmission
des messages des interleukines, des interfťrons et autres cytokines en
gťnťral mais aussi de l'hormone de croissance et de la prolactine par exemple.
Les rťcepteurs RCTK exercent leurs effets par l'intermťdiaire de protťines cytoplasmiques qui leurs sont associťs et qui possŤdent une activitť tyrosine kinase. Ils sont constituťs de chaÓnes polypeptidiques n'ayant qu'un seul segment transmembranaire. Leur activation par fixation du ligand induit en gťnťral une dimŤrisation (parfois une oligomŤrisation) de chaÓnes induisant ŗ son tour une activation des tyrosines kinases associťes (figure 9-37).
Les types de tyrosines kinases associťes sont nombreuses et varient avec le rťcepteur considťrť. Les plus connues ŗ l'heure actuelle sont JACK 1, 2 et 3 ou encore TYK2. L'activitť de ces enzymes induit la phosphorylation d'autres protťines qui leurs servent de substrat. Parmi celles-ci, citons les STATs (Signal Transducer and Activator of Transcription) qui sont des facteurs importants dans les rťgulations de transcription impliquťes dans les effets physiologiques de nombreuses sťmiomolťcules (figure 9-37).
Figure 9-37 : Activation et transmission du signal au niveau des rťcepteurs RCTK. La liaison du ligand (L) induit une dimŤrisation d'un rťcepteur (R) avec activation d'une protťine associťe (PA, ici JACK). La tyrosine kinase est ainsi phosphorylťe et peut phosphoryler une protťine substrat (ici STAT).
4.3.4. Rťcepteurs ŗ activitť
tyrosine kinase - SystŤme RATK
Dans ce systŤme, la fixation du ligand induit une activitť tyrosine kinase
dans le domaine intracellulaire du rťcepteur avec pour consťquence une
autophosphorylation des tyrosines de la chaÓne. Diffťrentes protťines-cibles
pourraient alors Ítre phosphorylťes et induiraient une activitť biologique (figure 9-38).
Font partie de ce systŤme les rťcepteurs activťs notamment par l'insuline et une sťrie de facteurs de croissance (EGF : epithelial growth factor Ė FGF : fibroblast growth factor Ė NGF : nerve growth factor Ė PDGF : platelet derived growth factor Ė VEGF : vascular endothelial growth factor).
Les rťcepteurs RATK sont constituťs d'une seule chaÓne polypeptidique transmembranaire (pour les facteurs de croissance) ou de deux chaÓnes (pour l'insuline). L'extrťmitť N-terminale, extracellulaire, de la chaÓne est en gťnťral glycosylťe et peut comporter plusieurs domaines diffťrents. L'extrťmitť C-terminale est intracellulaire et porte l'activitť catalytique.
La fixation du ligand entraÓne une dimŤrisation des rťcepteurs monomŤriques. Cette ťtape n'intervient pas au niveau des rťcepteurs dťjŗ dimŤriques comme celui de l'insuline par exemple. La liaison ligand-rťcepteur induit un changement de structure des domaines cytoplasmiques qui permet l'activation des tyrosines kinases. Il en rťsulte une phosphorylation croisťe des deux chaÓnes, le domaine cytoplasmique d'une chaÓne phosphorylant l'autre. Ces phosphorylations induisent l'apparition de sites ŗ haute affinitť pour des protťines substrats. Elles sont alors phosphorylťes ce qui active leurs propres activitťs biologiques (cfr. exemples dans la section 4.4).
L'inactivation du systŤme fait semble-t-il intervenir une internalisation par endocytose. Les rťcepteurs sont ainsi incorporťs dans des vťsicules d'oý ils peuvent Ítre recyclťs aprŤs dissociation du ligand.
Figure 9-38 : Activation et transmission du signal au niveau des rťcepteurs RATK. La liaison du ligand (L) au rťcepteur (R) conduit ŗ une dimŤrisation dans le cas des rťcepteurs monomŤriques (celui du facteur de croissance par exemple). Certains rťcepteurs sont au dťpart dimŤriques ; celui de l'insuline comporte deux sous-unitťs a extracellulaires, liťes ŗ deux sous-unitťs b transmembranaires par des ponts disulfures. L'autophosphorylation croisťe des domaines catalytiques intracellulaires permet l'activation par phosphorylation de protťines substrats (PS) ayant diffť-rentes activitťs biologiques. Dťtails dans le texte.
4.3.5. Rťcepteurs ŗ activitť
autre que tyrosine kinase
Sont repris dans ce cadre gťnťral les rťcepteurs ŗ activitť
sťrine/thrťonine kinase comme ceux des TGF (transforming growth factors), les
rťcepteurs ŗ activitť histidine kinase ainsi que les rťcepteurs ŗ activitť
tyrosine phosphatase de type CD. Ces derniŤres paraissent intervenir
essentiellement dans la transmission de signaux immunitaires dans les
lymphocytes B et T. Ils seront considťrťs briŤvement au chapitre 10.
Le mťcanisme d'activation des rťcepteurs des TGF est complexe et loin d'Ítre clairement dťfini. Il ferait intervenir en cascade 3 types de rťcepteurs diffťrents (fig.9-39). Le ligand se fixerait d'abord sur un rťcepteur de type III, ce qui permettrait son transfert successivement sur un rťcepteur de type II puis sur un rťcepteur de type I possťdant une activitť kinasique mais incapable de fixer directement le ligand. L'activitť kinasique serait induite par phosphorylation de rťsidus sťrines du rťcepteur I par le rťcepteur II suite ŗ un phťnomŤne de recrutement. L'activitť kinasique du rťcepteur I serait responsable de la phosphorylation de constituants cytoplasmiques ŗ effets biologiques (fig. 9-39).
Figure 9-39 : Mťcanisme possible d'activation et de transmission du signal au niveau des rťcepteurs ŗ activitť sťrine/thrťonine kinase. Dťtails dans le texte.
| |
|