Il ressort clairement de ce qui précède que le contrôle de l'osmolarité sanguine fait intervenir différents mécanismes impliquant des mouvements d'eau et d'effecteurs osmotiques au niveau de différents épithélia.
Les effecteurs osmotiques majeurs sont essentiellement les ions inorganiques Na+, K+ et Cl- ainsi que quelques solutés organiques : urée et différents composés aminés. Le contrôle des concentrations de ces composés met en œuvre des mécanismes de transport transépithéliaux. De tels mécanismes sont également impliqués dans des processus autre que l'osmorégulation comme par exemple : (cfr. chapitre 5) production d'acide au niveau stomacal, transfert au niveau de la barrière intestinale de sucres simples, d'acides aminés ou d'acides gras provenant de la digestion de différents aliments, sécrétion des glandes salivaires, du pancréas, de la vésicule biliaire, etc… ou encore (cfr. chapitre 12) sécrétion des glandes sudoripares.
Outre leur rôle majeur dans le contrôle de l'osmolarité sanguine, les mouvements d'eau sont d'importance capitale dans les processus de sécrétion de fluides au niveau de différentes glandes et organes ainsi que dans les processus d'absorption d'eau au niveau intestinal et rénal ou encore dans les processus de contrôle du volume et de l'osmolarité intracellulaire.
Comme nous le verrons par ailleurs dans la suite, les mouvements de certains ions sont à la base des propriétés électriques des membranes ; propriétés qui seront exploitées dans différents processus tels que la génération de potentiels d'action intervenant dans les interactions nerveuses (cfr. chapitre 8) ou encore la contraction musculaire (cfr. chapitre 7).
L'étude des mouvements de substances à travers les membranes biologiques revêt donc un intérêt majeur pour le physiologiste. Elle a d'ailleurs fait l'objet d'un nombre croissant de travaux de recherche et de publications scientifiques au cours des trente dernières années.
Une étude détaillée de ces mécanismes sort du cadre de ce traité, nous nous contenterons ici de présenter brièvement quelques-uns des processus de base qui sous-tendent les activités physiologiques qui y sont envisagées.
L'essentiel des mouvements de substances qui nous intéressent ici ont lieu à travers des membranes biologiques, soit membranes plasmiques cellulaires, soit encore membranes épithéliales ou endothéliales constituées essentiellement de cellules associées.
Les premiers travaux considérant l'existence de membranes entourant les cellules furent entrepris au milieu du 19ème siècle par les botanistes Nägeli et Pfeffer. Ce n'est cependant que relativement récemment, au cours des trente dernières années, que nombre de biologistes ont considéré les membranes plasmiques comme des structures aussi importantes et essentielles à la vie cellulaire que les autres : noyau, mitochondrie ou reticulum endoplasmique par exemple. Ces membranes sont en fait des systèmes biologiques très complexes constituant des barrières dynamiques sélectives entre deux compartiments très différents l'un de l'autre : le milieu extracellulaire d'une part (milieu OUT) et le milieu intracellulaire de l'autre (milieu IN). Elles comportent, enchâssées dans une base de nature lipidique, des structures, essentiellement protéiques, qui sont à la base de leurs activités ( figure 1-24). On y retrouve ainsi des canaux, des transporteurs, des récepteurs et des structures de liaison.
Les récepteurs et les structures de liaison vont intervenir dans tous les processus d'adhésion, de reconnaissance cellulaire, de structuration tissulaire ou encore de communication d'information entre cellules ou entre cellules et milieu environnant.
Les canaux et les transporteurs vont intervenir dans les mouvements d'ions, d’eau et de molécules auxquels sont par exemple associés : les apports d'éléments nutritifs et la sortie de produits du catabolisme ou d'éléments nouvellement synthétisés, les phénomènes bioélectriques et en particulier la génération des potentiels d'action et la communication nerveuse, le contrôle du volume cellulaire et le contrôle de la contraction musculaire.
Figure 1-24 : Eléments et rôles des membranes cellulaires (plasmiques). Les systèmes effecteurs répartis sur la membrane sont enchâssés ou ancrés dans une bicouche lipidique.
Les membranes de cellules constituent, comme les membranes plasmiques cellulaires, des barrières dynamiques entre deux compartiments qui sont ici deux milieux extracellulaires. Dans le cas des membranes épithéliales, le milieu IN fait classiquement référence au milieu intérieur, liquide sanguin le plus souvent. La face de l'épithélium tournée de ce côté portera dès lors souvent le nom de face séreuse. On l'appellera également face basale ou interne. Le côté OUT, à l'inverse, fera référence à la face apicale ou externe de l'épithélium. Cette face sera encore appelée luminale lorsque l'épithélium sépare la lumière d'un organe (intestin, néphron par exemple) du milieu intérieur (figure 1-25).
Les membranes endothéliales pour leur part, séparent généralement le sang d'un autre liquide interne : liquide interstitiel pour l'endothélium vasculaire - liquide cérébro-spinal pour la barrière hémato-encéphalique par exemple.
Figure 1-25 : Eléments et rôles des membranes de cellules (épithélia - endothélia). Les messages reçus et les mouvements d'entrée et de sortie concernent souvent des effecteurs différents sur les deux faces. Les mouvements transépithéliaux peuvent s'effectuer à travers les cellules (ou uniquement certains types cellulaires) mais aussi à travers les espaces intercellulaires.
Généralement les éléments structuraux responsables des différentes activités sont organisés de façon asymétrique, la face externe ayant des caractéristiques spécifiques, avec canaux, transporteurs et récepteurs différents de ceux présents sur la face interne. Les membranes plasmiques des cellules constitutives peuvent d'autre part former des espaces extracellulaires très complexes formant des systèmes particuliers dont il faudrait pouvoir tenir compte dans l'étude des mouvements transmembranaires. Ce type d’organisation spécifique est particulièrement frappante au niveau des cellules des épithélia transporteurs dont les membranes plasmiques forment par invagination des réseaux extrêmement complexes (figure 1-26).
Les épithélia et endothélia sont constitués de cellules associées les unes aux autres avec plus ou moins de cohésion. Les mouvements transmembranaires pourront dès lors s'effectuer via les cellules et/ou les espaces intercellulaires, en fonction, notamment, de la cohésion plus ou moins importante du tissu considéré.
Figure 1-26 : Deux exemples de structures épithéliales mettant particulièrement bien en évidence la complexité des espaces extracellulaires : le tubule contourné proximal du rein de mammifère (A - B) et la branchie postérieure du crabe chinois Eriocheir sinensis (C - D). Les deux faces de ces épithélia sont largement plissées, permettant un accroissement de surface bien adapté à leur rôle dans différents mécanismes de transport (NaCl pour la branchie - NaCl, eau et solutés organiques pour le tubule proximal). Au niveau du tubule proximal, les plissements apicaux forment des microvillosités similaires à celles rencontrées au niveau de la bordure en brosse de l'intestin grêle (voir chapitre 5, 4.2). Comme au niveau de l'intestin également, les cellules sont associées entre elles par des complexes jonctionnels apicaux (flèches) formant des jonctions serrées bien adaptées à la fonction de réabsorption d'eau de ces tissus. Ces jonctions serrées sont toutefois moins élaborées que la zonula occludens des cellules intestinales suggérant qu'elles puissent être moins imperméables que ces dernières. Les importants plissements baso-latéraux sont bourrés de mitochondries qui assurent l'apport énergétique nécessaire au fonctionnement des Na+/K+ATPases de transport. Les plissements apicaux forment par ailleurs avec les matrices extracellulaires (glycocalyx) produites par les cellules un milieu particulier qui pourrait jouer un rôle spécifique dans certains systèmes de transport. Bien que ce rôle reste encore à définir, on peut par exemple montrer que la structuration des pôles apicaux et baso-latéraux de la branchie ainsi que sa capacité à transporter les ions varient largement chez le crabe E.sinensis en eau douce (figure C : pompage actif de NaCl important) ou en eau de mer (figure D : pas de pompage actif de NaCl). Vues en microscopie électronique : A : d'après Weiss 1988, modifié ; B : d'après Cross et Mercer 1995, modifié ; C et D : d'après Péqueux et Gilles, originaux ; les deux vues sont au même grossissement. (Cut : cuticule, Lum : lumière, M : mitochondrie, S : sang).
Certains épithélia peuvent par ailleurs comporter différents types de cellules en une ou plusieurs couches, ces cellules pouvant jouer des rôles tout à fait différents dans les processus de transport. Dans certains cas, les cellules adjacentes sont de plus reliées entre elles par des jonctions lacunaires (ou communicantes) mettant en connexion le liquide intracellulaire d'une cellule avec celui de ses voisines (figure 1-27).
Figure 1-27 : Représentation schématique d'une jonction lacunaire (communicante - gap junction) entre deux cellules. Ces structures, très nombreuses dans certains tissus, permettent le passage d'une cellule à l'autre de solutés allant jusqu'à un poids moléculaire de l'ordre de 1000 à 1500 D (ions, éléments nutritifs, etc...). Elles forment en fait un canal constitué par l'association de deux éléments transmembranaires (connexons) formant un pore de ±1,5 nm entouré de 6 sous-unités identiques de connexine. Les connexons sont généralement groupés en plaques plus ou moins larges (fascia) au niveau desquels l'espace intercellulaire est très réduit (2 à 3 nm au lieu de 10 à 15 nm).
Les membranes épithéliales sont donc des structures beaucoup plus complexes que les membranes plasmiques. Elles présentent toujours un nombre de compartiments plus élevé que ces dernières ; chacun de ceux-ci pouvant avoir un rôle particulier à jouer dans un processus de transport (figure 1-28). L'organisation de ces différents compartiments les uns par rapport aux autres ainsi que la présence de différents transporteurs, canaux ou récepteurs sur les faces apicales et latéro-basales permet par ailleurs une grande variabilité dans la structure des épithélia et une grande diversité de fonctions : iono et osmorégulation sanguine, contrôle de la volémie, sécrétion de fluides, transport d'éléments nutritifs ou de déchets ...
Figure 1-28 : Membranes cellulaires et membranes de cellules - complexité très différente des deux systèmes dans le cadre des mouvements transmembranaires. Au niveau de la cellule, le passage se fait entre les compartiments IN et OUT à travers la membrane plasmique considérée comme un compartiment homogène unique, les systèmes effecteurs étant répartis sur toute la surface. Au niveau des membranes de cellules, les mouvements transmembranaires peuvent s'effectuer en passant successivement, dans le cas d'un mouvement OUT - IN, par une série de 9 compartiments comprenant ou pouvant comprendre un espace extracellulaire externe spécifique (2), une membrane plasmique apicale (3), un espace intracellulaire (4), une membrane plasmique basale (5) et, éventuellement, un espace extracellulaire interne spécifique (6). Des mouvements transépithéliaux peuvent également avoir lieu via les espaces intercellulaires (9 et 9'), ceux-ci étant parfois structurés de façon particulière comme dans le cas des jonctions serrées de l'intestin par exemple (9'). Certains mouvements peuvent par ailleurs s'effectuer entre les cellules, à travers des jonctions lacunaires de connexion (10). Les systèmes effecteurs sont répartis différemment et spécifiquement sur les deux faces de la membrane. Différents épithélia peuvent par ailleurs comporter différents types cellulaires jouant des rôles physiologiques tout à fait différents. (Détails dans le texte).
L'organisation des membranes plasmiques intervient de façon majeure dans les mouvements transmembranaires, qu'ils s'effectuent au niveau d'une cellule ou d'un épithélium. Elles sont constituées d'une double couche de lipides, organisés en deux feuillets continus dans lesquels sont incorporés des éléments de nature protéiques, glyco-protéiques, oligo-saccharidiques ou glycolipidiques représentant les récepteurs, canaux, transporteurs et autres éléments "actifs" des membranes.
Le premier modèle de membrane plasmique qui correspondait le mieux aux observations de microscopie électronique fut celui que Davson et Danielli proposèrent dès 1935. La membrane apparaissait alors comme une bicouche lipidique de 5 à 6 nm d'épaisseur (50-60 Å) avec des éléments protéiques "associés" internes et externes. Si les concepts concernant l'arrangement de ces éléments ont largement évolué depuis (voir 5.3.2 ci-après), l'idée de la bicouche de lipides a reçu un agrément universel. L’organisation des éléments "associés", protéiques et autres, a été mise en évidence par différentes techniques dont celle du cryodécapage qui permet la séparation de la bicouche en ses deux feuillets, l’un externe et l’autre interne (figure 1-29B). On a ainsi pu mettre en évidence la présence d’éléments "associés" externes, internes (dits extrinsèques) ou encore traversant la bicouche (dit intrinsèque). D’autres techniques ont par ailleurs montré que ces éléments étaient sujet à mouvement, regroupement et remplacement. On est ainsi arrivé au modèle dit en mosaïque fluide présenté dans la figure 1-29C.
Les lipides des membranes plasmiques des cellules animales appartiennent principalement au groupe des phospholipides (figure 1-30). On y trouve également des sphingolipides et du cholestérol en quantités variables selon les types cellulaires. L'ensemble des lipides intervient en général pour quelque 50% dans la composition chimique globale des membranes (voir plus loin tableau 1-9).
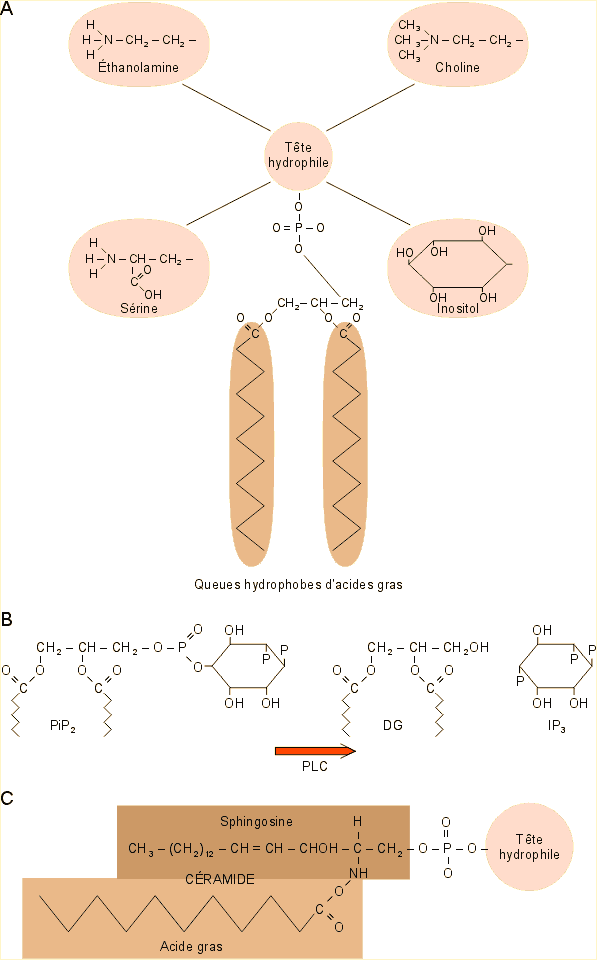
Tous les phospholipides sont dérivés de l'acide phosphatidique, structure correspondant à un glycérol phosphorylé, auquel sont ajoutés d'un côté deux "queues" hydrophobes d'acides gras et de l'autre une "tête" hydrophile. L'ensemble constitue une structure dite amphiphile avec une partie polaire et une partie apolaire situées dans deux régions distinctes de l'espace. C'est cette structure qui va conférer à la membrane l'essentiel de ses caractéristiques de base. En milieu aqueux en effet les lipides s'organisent automatiquement en bicouche ; organisation qui permet de minimiser au mieux les interactions entre les parties hydrophobes, tournées vers l'intérieur, et les parties hydrophiles, tournées vers l'extérieur (voir b. ci-après).
La nature des "têtes" et des "queues" est éminemment variable. Les têtes que l'on trouve le plus souvent sont la choline, l'éthanolamine, la sérine et l'inositol (figure 1-30A). Les phospholipides formés sur cette base formeront respectivement les groupes de la phosphatidylcholine (PC), de la phosphoéthanolamine, de la phosphosérine et du phosphatidylinositol (PI). On trouve dans ce dernier groupe un lipide très minoritaire, le phosphatidylinositol biphosphate (PiP2) qui joue en fait un rôle particulier dans la communication chimique. Son hydrolyse par la phospholipase C conduit à la formation d'un diglycéride, le diacylglycérol (DG) et d'inositoltriphosphate (IP3) qui vont intervenir dans un système de signalisation cellulaire (voir B figure 1-30 et chapitre 9).
À côté des phospholipides, on trouve également des sphingolipides dont le constituant de base est la sphingosine, un alcool aminé à longue chaîne. Son amidification par une "queue" d'acide gras hydrophobe forme une céramide à laquelle peut être ajoutée une "tête" hydrophile par l'intermédiaire d'un groupement phosphate comme dans le cas des phospholipides. Ainsi, dans la sphingomyéline, composante importante des gaines de différents nerfs (voir chapitre 8), la céramide est prolongée par une tête de phosphorylcholine (C figure 1-30).
Figure 1-30 : Les lipides des membranes plasmiques : A : phospholipides avec 4 des têtes majeures - B : Formation de deux molécules de signalisation intracellulaire importante (IP3 et DG) par hydrolyse de Pi P2 - C: sphingophospholipides. Détails dans le texte.
Les acides gras formant les queues hydrophobes sont nombreux. On en a caractérisé plus de 900 à l'heure actuelle. Leur longueur varie entre 12 et 24 atomes de carbone, ils peuvent être saturés ou insaturés (avec un nombre de doubles liaisons allant de 1 à 6) et parfois branchés.
Le nombre élevé d'acides gras utilisables, couplé à l'utilisation des différentes têtes possibles, fait que le nombre de phospholipides théoriquement possible est en principe très important. En pratique, on n'en trouve guère plus d'une cinquantaine au niveau d'un type de membrane donné et le plus souvent, il n'y en a que quelques-uns qui soient présents en quantités importantes (voir tableau 1-9).
TABLEAU 1-9
Composition en lipides majeurs de préparations de membranes plasmiques
de différents tissus du rat
|
Foie |
Globule rouge |
Axones myélinisés |
|
| Protéines | 44 | 49 | 18 |
| Glucides | 4 | 8 | 3 |
| Lipides | 52 | 43 | 79 |
| dont : | |||
| Cholestérol | 30 | 24 | 22 |
| PC | 18 | 31 | 11 |
| PE | 11 | 15 | 14 |
| PI | 4 | 2 | – |
| PS | 9 | 7 | 7 |
| SM | 14 | 8,5 | 6 |
| PA | 1 | 0,1 | – |
| Glycolipides | – | 3 | 12 |
Les caractéristiques générales des lipides impliqués confèrent à la membrane l'essentiel de ces caractéristiques de base. Nous n'en considérerons ici très brièvement que quelques unes.
1) Comme nous l'avons déjà signalé, la nature amphiphilique de ces molécules, avec leur région hydrophile et hydrophobe en opposition, est à la base de la formation en bicouche, structure permettant de minimiser au mieux les interactions entre les régions. Cette structure a par ailleurs la propriété de se refermer sur elle-même pouvant ainsi former des vésicules de plus ou moins grandes dimensions et donc des cellules. Dans ce contexte, les cellules n'existent que dans la mesure où il y a des membranes lipidiques. Le premier rôle des lipides est donc un rôle de compartimentation permettant l'organisation de milieux aqueux différents de part et d'autre de la membrane.
Cette propriété à former des bicouches linéaires s'enroulant en vésicules a été largement utilisée en recherche. On peut ainsi dans certaines conditions générer des "ghosts" (fantômes) de cellules en éliminant par extrusion le cytoplasme et tous les éléments figurés intracellulaires. La bicouche, rompue lors de la préparation, se referme automatiquement, rendant une membrane plasmique dont on peut étudier les caractéristiques de transport sans influence des structures intracellulaires. On peut également préparer des "liposomes" de type vésiculaire et des monocouches ou bicouches linéaires (black membranes) après dissolution des lipides dans un solvant approprié, concentration et remise en suspension dans un solvant aqueux (figure 1-31).
La propriété qu'ont les bicouches de s'organiser en vésicules est utilisée pour fabriquer des liposomes dont une application majeure actuelle est l'encapsulage de médicaments ou de produits cosmétiques. En fonction du type de préparation, les liposomes peuvent être multilamellaires, présentant plusieurs bicouches organisés de façon concentrique, ou bien unilamellaires de petite ou grande taille. Ces derniers sont obtenus par formation d'une monocouche de lipides dissous dans l'éther, à l'interface éther-eau. Le système est ensuite émulsionné par ultrasons pour obtenir des monocouches micellaires contenant de l'eau. L'évaporation lente de l'éther provoque alors une réorganisation des micelles avec formation de liposomes de grande taille qui peuvent être placés en milieu aqueux. Cette dernière préparation convient par ailleurs très bien aux études physiologiques de flux. Il en va de même pour les bicouches planes ("black" membranes des anglo-saxons) que l'on obtient en couchant un film de lipides dissous dans un solvant organique non nuisible à l'eau sur l'orifice circulaire d'une paroi séparant deux compartiments aqueux. Il est également possible d'insérer des protéines de membranes dans les bicouches; ce qui a permis une certaine approche de l'étude de leurs propriétés. Les caractéristiques de perméabilité et de transport pourront également être étudiées sur des "fantômes" ("ghosts") de cellules (globules rouges annuclés de mammifères essentiellement) obtenus par lyse en milieu hypoosmotique des éléments intracellulaires. Une fois ces éléments éliminés, les érythrocytes sont remis en milieu isotonique dans lequel les membranes se réparent donnant ainsi des préparations de type bicouche contenant tous les éléments protéiques constitutifs de départ.
Les vésicules et surtout les bicouches se sont révélées intéressantes pour étudier la perméabilité à différents ions ou solutés de membranes purement lipidiques, sans constituants protéiques (voir 5.4 ci-après). Les vésicules sont par ailleurs à la base de la préparation des liposomes, structures rondes et closes, le plus souvent en bicouches concentriques ayant trouvé des applications en cosmétologie, en pharmacologie et en médecine. Il est en effet possible "d'encapsuler" dans ces liposomes différents agents, rendant ainsi leur utilisation plus aisée ou plus efficace. Les monocouches ont trouvé des applications dans l'étude des interactions lipides/protéines et dans l’étude des structures protéiques en cristaux bidimensionnels.
2) Les acides gras constituant la partie centrale des bicouches leurs confèrent les caractéristiques d'un liquide huileux. Cette propriété est importante dans le cadre des phénomènes de diffusion à travers les membranes plasmiques comme nous le verrons par la suite (voir 5.4 ci-après). Elle intervient également dans la fluidité plus ou moins grande des membranes. Cette fluidité paraît importante physiologiquement dans la mesure où elle conditionne les possibilités d'insertion et de mouvements des divers éléments constitutifs, protéines notamment. La viscosité des membranes plasmiques est, comme celle de tout mélange huileux, fonction de la conformation et de la nature des chaînes des acides gras constitutifs. Elle est par ailleurs variable avec la température ; une diminution de celle-ci induisant une augmentation de viscosité qui peut s'avérer néfaste au fonctionnement correct des membranes. On remarque dans ce cadre que les lipides des membranes plasmiques de cellules d'animaux pœcilothermes de régions froides ont des chaînes d'acides gras présentant un degré d'insaturation plus élevé que ceux d'animaux de régions chaudes (voir chapitre 12). Le nombre de doubles liaisons détermine en effet la température de fusion et la fluidité plus ou moins grande des acides gras et donc des membranes plasmiques. L'augmentation du nombre d’acides gras insaturés au sein des membranes plasmiques constitue donc une adaptation intervenant chez les pœcilothermes de régions froides dans le maintien des fonctions normales des cellules.
3) Les lipides de membranes sont animés de mouvements divers, plus ou moins importants et rapides en relation avec la fluidité des couches huileuses d'acides gras. On peut ainsi observer des mouvements de balancier des chaînes hydrocarbonées et de rotation ou de translation des têtes (figure 1-32). Ces mouvements sont importants surtout dans la mesure où ils facilitent les insertions et donc le renouvellement ainsi que les mouvements de diffusion latérale des protéines dans les membranes (voir 4 ci-après).
Figure 1-32 : Mouvement des phospholipides de membrane et des éléments protéiques associés (détails dans le texte).
4) Les membranes plasmiques apparaissent à l'heure actuelle comme des associations spécifiques de quelque 50 lipides qui ne sont pas distribués au hasard, mais en microdomaines dans lesquels vont, semble-t-il, pouvoir s'insérer différents types de protéines. Celles-ci, de même que les lipides des bicouches, présentent des mouvements de diffusion latérale leurs permettant de se déplacer lentement dans un domaine et même d'un domaine à l'autre. Certains domaines pourraient aussi permettre des déplacements latéraux plus faciles Les caractéristiques de transport, d'activité endocytaire ou de perméabilité pourraient dès lors ne pas être distribuées uniformément sur la surface d'une cellule.
Il apparaît également que les lipides ne sont pas distribués de façon symétrique dans les bicouches. Ainsi il y a dans les membranes de globules rouges humains plus de sphingomyéline et de phosphatidylcholine du côté extérieur et plus de phosphatidyléthanolamine et de phosphatydylsérine du côté intérieur.
Les mécanismes assurant la sélection et la bonne insertion des phospholipides dans les microdomaines ou sur les faces de la membrane restent à l'heure actuelle inconnus. Les différents phospholipides et céramides sont synthétisés au niveau de la surface cytosolique du reticulum endoplasmique. Les céramides sont alors converties en sphingolipides au niveau du Golgi. Les lipides formés sont alors véhiculés jusqu'à la membrane par un système de trafic vésiculaire. Comment s'effectue le choix des lipides à véhiculer et comment sont-ils dirigés vers la zone de membrane correcte, restent pour l'instant des questions sans réponses précises. Ce trafic peut être très rapide ; les membranes plasmiques apparaissent dès lors comme des assemblages dynamiques capables d'adaptations rapides permettant une régulation d'activité en fonction de différents paramètres : hormones, modifications des conditions des milieux extérieurs ou intérieurs par exemple.
Les éléments concernant les microdomaines et la mobilité des protéines en leur sein sont relativement récents et leur incidence physiologique réelle au niveau d'une cellule isolée n'est pas encore très claire. Au niveau des membranes de cellules, l'organisation des microdomaines conduit en plus à une spécialisation des faces apicales et basales ; chacune d'elles présente en effet des activités spécifiques comme nous le verrons par la suite (voir 5.4). La cellule nerveuse est également un exemple de système ou les microdomaines ont permis l'organisation de structures particulaires tels qu'axones avec canaux ioniques ou régions transmettrices avec vésicules à neurotransmetteurs et récepteurs par exemple (voir chapitre 8).
De nombreuses structures, de nature essentiellement protéiques sont insérées dans les bicouches membranaires. Elles y jouent un rôle physiologique majeur, étant responsables pour l'essentiel des activités caractéristiques des membranes : transport, reconnaissance, communication, liaison.
Dans le schéma actuel, dit de "mosaïque fluide" de la membrane, on distingue des protéines intrinsèques et extrinsèques ou encore intégrales et périphériques (figure 1-29) (Singer et Nicholson, Science 175, 720, 1972).
Les protéines intrinsèques forment des structures transmembranaires avec une partie insérée dans la bicouche, une partie dépassant du côté externe et une autre du côté interne. La bicouche ayant un caractère huileux, les segments qui y sont insérés doivent présenter un degré hydrophobique suffisant. Ils prennent dès lors le plus souvent une structure a-hélicoïdale (figure 1-33). Cette structure permet en effet un maximum de liaisons entre les groupements C = O et N – H des acides aminés constitutifs qui ont un caractère polaire. Dans bon nombre de cas, les hélices transmembranaires sont en fait amphiphiles; bien qu'ayant un caractère général hydrophobe, elles comprennent un certain nombre d'acides aminés polaires déterminant des zones plus ou moins hydrophiles pouvant rendre compte d'une certaine orientation dans la membrane ou de l'organisation d'un canal central par exemple. Ce type de structure est généralement rencontrée dans les canaux et les transporteurs. On la rencontre aussi au niveau de certains récepteurs intervenant dans les processus de communication chimique (voir chapitre 9). Le nombre de segments a-hélicoïdaux des canaux et transporteurs varie en général entre 4 et 12 (voir 5.4 ci-après et aussi chapitre 8).
Ces structures forment des familles présentant un haut degré d'homologie. En parlant d'un transporteur donné on se réfère en fait à une famille de molécules présentant un même type d'activité dans des tissus ou des organismes différents. Nous considérerons certaines d'entre elles plus en détail en envisageant différents mécanismes de transport (voir 5.4 ci-après), ou encore la génération du potentiel d'action (voir chapitre 8).
Une autre organisation structurale présentant un certain degré d'hydrophobicité est la chaîne b ; les feuillets plissés de ces structures permettent en effet également des liaisons entre acides aminés, annihilant leur caractère polaire. L'organisation en feuillets b est courante au niveau de nombreuses protéines intrinsèques bactériennes comme les porines, on la rencontre beaucoup plus rarement au niveau des membranes plasmiques des cellules animales. Elle constitue vraisemblablement la structure d'un motif particulier des canaux Na+, K+ et Ca2+ potentiel-dépendant (voir figure 1-33, section 5.5, chapitres 7 et 8).
 |
 |
Figure 1-33 : Exemple de protéine intrinsèque : le canal K+ potentiel-dépendant : profil d'hydrophobicité et topologie probable d'une unité (A) - organisation de 4 unités formant le canal (B) et visualisation du canal dans la membrane (C). Les protéines intrinsèques des membranes plasmiques des cellules animales ont des segments transmembranaires essentiellement organisés en hélice a, structure thermodynamiquement favorable dans un milieu huileux. Le nombre probable de segments a-hélicoïdaux est déterminé à partir de la structure primaire des protéines en assignant à chaque acide aminé un indice d'hydrophobicité. En portant sur un graphique l'indice de l'acide aminé en fonction de sa position dans la séquence, on obtient un profil d'hydrophobicité qui fait apparaître autant de régions hydrophobes de 20 à 40 acides aminés qu'il y a d'hélices a. La structure plissée en chaîne b, qui permet également une insertion transmembranaire, paraît rare au niveau des membranes des cellules animales. La protéine prise ici comme exemple est une des rares à l'heure actuelle où l'on admet la présence de deux chaînes b à côté de 6 segments a-hélicoïdaux. Le canal complet est en fait constitué de l'association de 4 unités identiques. Il a une organisation générale voisine de celle du canal Na+ sensible au potentiel (voir 5.4.1 ci-après et chapitre 8).
Les protéines extrinsèques sont constituées de domaines essentiellement en contact avec un milieu polaire : milieu aqueux extra ou intracellulaire ou encore domine polaire des structures a -hélicoïdales dans le cas ou elles sont rattachées à celles-ci. Elles seront donc plus ou moins repliées sur elles-mêmes avec des chaînes latérales plus hydrophiles vers l'extérieur et des chaînes latérales plus hydrophobes vers l'intérieur.
La plupart de ces protéines sont associées à un segment hydrophobe permettant leur insertion dans la partie huileuse des bicouches (figure 1-34). Il s'agit le plus souvent d'associations covalentes avec un acide gras (myristique ou palmitique), un lipide (glycosylphosphatidyl-éthalanolamine : GPI) ou un résidu isoprényl (farsényl ou géranylgéranyl).
Les protéines myristoylées, palmitoylées ou isoprénylées sont toujours en contact avec le milieu intracellulaire. Les liaisons à la membrane par myristoylation ou palmitoylation sont relativement faibles et les protéines peuvent dès lors participer facilement à des séquences dans lesquelles elles s'attachent et se détachent de la membrane suite à phosphorylation. Elles sont dès lors souvent impliquées dans des processus de signalisation cellulaire. Ainsi, MARCKS est une protéine myristoylée apparaissant comme un substrat de la PKC et les sous-unités labiles (a ) de pratiquement toutes les protéines G sont palmitoylées (voir chapitre 9). Dans l'isoprénylation, la protéine est attachée par une cystéine terminale qui est carboxyméthylée. Ceci confère aux protéines un caractère hydrophobique plus marqué que la myristoylation ou la palmytoylation. Les protéines formeront donc des complexes membranaires plus stables. Ainsi, les sous-unités stables (g ) des protéines G sont isoprénylées (voir chapitre 9).
Figure 1-34 : Organisation générale des protéines extrinsèques montrant différents types d'ancrage dans la bicouche. Attaches N-myristoylglycine, S-palmytoylcystéine ou S-farnésyl et S-géranylgé-ranylcystéine (isoprénylées) pour le milieu intracellulaire. Attache C-GPI pour le milieu extracellulaire. La partie du motif GPI servant à l'ancrage membranaire est un phosphatidyl-inositol (PI). La partie C terminale de la protéine y est rattachée par l'intermédiaire d'une phosphoéthanolamine (PE) suivie de trois mannoses (Man) et d'une glucosamine (Gln) : les protéines isoprénylées sont rattachées par une cystéine carboxyméthylée.
Les protéines ancrées au GPI sont par contre localisées du côté extérieur ; elles vont donc jouer un rôle dans différents phénomènes de signalisation avec des éléments du milieu extérieur mais aussi d'adhésion et de reconnaissance cellulaire. Ainsi, la fibronectine qui joue un rôle particulièrement important dans la structuration en tissus par ces possibilités de rattachement aux protéines extracellulaires, collagène notamment.
Les protéines ancrées sur la face externe de la membrane forment avec leurs chaînes latérales et les domaines extracellulaires des protéines intrinsèques une sorte de "feutrage" auquel peuvent s'associer des productions cellulaires particulières comme du collagène ou du mucus par exemple. Ce revêtement (cell coat), appelé parfois glycocalyx, joue un rôle important dans les processus de reconnaissance. Il représente par ailleurs une couche de charge générale négative qui peut intervenir dans les mouvements de substances chargées à travers les membranes. Ainsi, il semble que le glycocalyx des cellules musculaires cardiaques est capable de "piéger" une grande quantité d'ions Ca2+ ; un processus qui pourrait faciliter la pénétration de l'ion au sein des cellules déclenchant ainsi leur contraction (voir chapitre 7). Le glycocalyx est particulièrement important au niveau de l'épithélium intestinal au niveau duquel il forme une couche épaisse au-dessus et entre les microvillosités (voir chapitre 5).
Les protéines ancrées sur la face interne ainsi que les parties intracellulaires des protéines intrinsèques sont le plus souvent associées directement à des protéines intracellulaires. Elles peuvent ainsi être en relation 1) avec des protéines faisant partie du système dit cytosquelettique, assurant un certain maintien de la forme des cellules (fibres d'actine, spectrine, ankirine, etc.) 2) avec des protéines assurant différentes activités métaboliques, ou encore 3) avec des protéines intervenant dans des systèmes de signalisation ou de trafic intracellulaire ( figure 1-35). Au niveau du globule rouge par exemple, on a pu montrer que les protéines membranaires intrinsèques associées au transport de Cl– sont en relation, par certaines parties de leur domaine intracellulaire, avec le cytosquelette d'une part, avec l'hémoglobine et avec certaines enzymes de la séquence glycolytique d'autre part (figure 1-35). Ces liaisons pourraient rendre compte des interactions métabolisme - volume cellulaire - transport observées dans certaines conditions au niveau de différents types cellulaires (voir 6 ci-après.).
Figure 1-35 : Représentations schématiques de différents systèmes de liaison entre protéines intrinsèques ou extrinsèques cytoplasmiques et systèmes effecteurs intracellulaires. Détails dans le texte.
Les relations entre protéines membranaires et intracellulaires peuvent être permanentes. Dans de nombreux cas, elles sont uniquement passagères, ne s'établissant que pour un temps très court dans le cadre d'une activité donnée. C'est notamment le cas dans différentes séquences de signalisation ou dans les séquences d'internalisation et d'adressage des vésicules endocytaires par exemple. Nous envisagerons certains de ces mécanismes en considérant plus spécifiquement ces problèmes (voir 5.4 ci-après, chapitres 8 et 9).
![]()
| |
|