4.1.1. Organisation générale
4.1.2. Les sécrétions de la muqueuse
4.1.3. Les mouvements de l'estomac
4.2.1. Organisation générale
4.2.2. Intestin grêle
4.2.3. Les sécrétions de la muqueuse
4.2.4. Gros intestin
4.2.5. Les mouvements intestinaux
4.3.1. Le pancréas
4.3.2. Le foie
Comme nous l'avons vu dans les pages qui précèdent, les structures essentiellement impliquées dans la digestion sont l'estomac et l'intestin. Les données concernant ces structures chez les invertébrés sont peu nombreuses, disparates et souvent préliminaires, nous nous concentrerons donc sur l'estomac et l'intestin des vertébrés.
4.1.1. Organisation générale
L'estomac des vertébrés inférieurs (poissons, amphibiens, reptiles) est la plupart
du temps tubulaire (figure 5-17). Il prend le plus souvent une forme
en j, en u ou en y. La forme droite, telle qu'on la retrouve chez certains poissons comme
la perche, est rare. Il est inexistant chez différents poissons (cyclostomes, dipneustes,
certains téléostéens). Chez d'autres, la portion inférieure (pylorique, voir plus
loin) est assortie de nombreux cæca ou appendices. La présence de ces cæca, dont le
nombre peut aller jusqu'à 1500, est une particularité des poissons. Ils ont, semble-t-il
une fonction glandulaire digestive et une fonction d'absorption. Les cæca s'abouchent en
fait au tube digestif au niveau du pylore à la limite entre l'estomac et l'intestin. Dans
la plupart des cas, ils sont localisés après la constriction pylorique, faisant donc
plus partie de l'intestin que de l'estomac quoique classiquement considérés avec ce
dernier.
Chez les oiseaux, l'estomac est partagé en deux parties distinctes par une constriction transversale (figure 5-17). La partie antérieure est glandulaire et secrète le suc gastrique, la partie postérieure est musculaire. Particulièrement bien différenciée chez les granivores et les herbivores, elle forme une poche, le gésier, qui assure un malaxage efficace des aliments très durs comme certaines graines ou parties ligneuses d'herbes sèches. Certaines espèces avalent de la pierraille ou du sable grossier qui, accumulé dans le gésier, aide au broyage mécanique. On trouve de ces "gastrolithes" également chez certains reptiles (crocodiles notamment). Il semble qu'ils étaient déjà présents chez certains dinosaures. La puissance de broyage et d'usure de l'estomac de certains oiseaux devient ainsi très grande. Une dinde est, parait-il, capable d'écraser dans son gésier un bout de tuyau de plomberie pouvant résister à une pression de 40 kg/cm2. Dans certaines régions, on utilise régulièrement des oiseaux (oies, autruches) pour donner rapidement une apparence antique à des copies récentes de statuettes ou de monnaies anciennes.
Figure 5-17: Estomac unilobulaire de différentes espèces non mammaliennes. Remarquez les différents types d'appendices pyloriques chez les poissons (saumon, truite, esturgeon). O: œsophage, G: région gastrique (fundus + corps), P: région pylorique, I: intestin.
Chez la plupart des mammifères, l'estomac reste simple, unilobulaire ou "monogastrique". Chez les ruminants ainsi que chez certains autres herbivores et différentes espèces aquatiques (dauphins, baleines, dugongs), il se divise en poches, essentiellement d'origine œsophagienne (figure 5-18). Chez les herbivores, ces poches interviennent dans le processus de digestion du matériel ligneux (voir 3.3.2). On ignore le rôle de ces poches chez les autres espèces. Elles pourraient servir d'espace de stockage, augmentant ainsi la durée du transit et donc l'efficacité de la digestion.
L'estomac est alimenté par l'œsophage qui s'y abouche chez l'homme au niveau d'une région dite cardiaque (figure 5-19). L'œsophage est fermé à ce niveau par un sphincter et une plicature de la couche musculaire œsophagienne. L'extrémité inférieure de l'estomac constitue la région pylorique qui s'ouvre sur le duodénum par l'orifice pylorique: celui-ci est normalement fermé par un sphincter qui régit l'évacuation gastrique. On définit également une région fundique, représentant chez l'homme la tubérosité en forme de dôme qui fait saillie à côté du cardia et un corps qui est la portion médiane de l'estomac. La structure microscopique du
corps est semblable à celle de la région fundique; on les regroupera ici sous le terme de région gastrique (d'après Stevens, 1988). Ces trois régions (cardiaque, gastrique et pylorique) ont des activités sécrétrices. Chez bon nombre d'espèces, on distinguera encore une région squameuse de structure proche de celle de l'œsophage. Ces différentes régions occupent des territoires de l'estomac de surface éminemment variable en fonction des espèces (figures 5-17 et 5-18). On remarquera cependant l'importance de la région gastrique chez les carnivores et les insectivores par rapport aux herbivores.
Figure 5-18: Estomac de différents mammifères, y compris ruminants. Remarquez la variabilité de l'importance de la région gastrique, notamment des herbivores (marsupiaux, rongeurs, artio et perissodactyles) par rapport aux autres. Les proportions par rapport aux tailles réelles ne sont pas respectées.
4.1.2. Les sécrétions de la muqueuse
Le revêtement de base de la muqueuse de l'estomac est composé de cellules
épithéliales fabriquant un mucus protecteur sous forme d'un gel alcalin. Toute
stimulation mécanique superficielle augmente la vitesse de formation du mucus. Le
parasympathique paraît également exercer une action stimulante alors que la gastrine
reste sans effet.
L'épithélium de surface s'invagine régulièrement pour former des favéoles au niveau desquels s'ouvrent des glandes. Les orifices de ces glandes constituent ce que l'on appelle les cryptes de l'estomac (plus ou moins 100 par mm2 chez l'homme, figure 5-19). Dans la région squameuse œsophagienne se trouve des glandes dites cardiaques œsophagiennes qui sécrètent essentiellement du mucus. La surface des autres régions est ponctuée par les orifices des glandes dites gastriques.
Les glandes gastriques sont constituées de différents types de cellules dont l'importance en nombre varie d'une région à l'autre, produisant des sécrétions différentes. On trouve essentiellement (figure 5-19) : 1) des cellules non différenciées qui vont servir à remplacer les cellules endommagées de la muqueuse superficielle. 2) Des cellules à mucus au niveau du collet. Ces cellules sécrètent un mucus différent de celui de l'épithélium superficiel et dont on ignore encore la fonction spécifique. 3) Des cellules pariétales, encore appelées bordantes ou oxyntiques, qui sécrètent les ions H+ et Cl-. 4) Des cellules principales qui élaborent les pepsinogènes. 5) Des cellules endocrines qui sécrètent notamment de la gastrine, de la sérotonine, de l'histamine, de la somatostatine. La gastrine et l'histamine vont stimuler la production de HCl des cellules pariétales alors que la sérotonine provoque la contraction des couches musculaires lisses entourant l'estomac. La somatostatine inhibe la production de gastrine alors que l'histamine l'active. La gastrine stimule également l'évacuation du contenu stomacal vers le duodénum alors qu'elle paraît inhiber le tonus de l'estomac proximal (voir 4.1.3 ci-après). Les cellules pariétales sécrètent par ailleurs un facteur dit intrinsèque requis pour l'absorption intestinale de la cyanocobalamine (vitamine B 12), nécessaire à la production de globules rouges matures. La production d'histamine, de sérotonine et de somatostatine paraît sous contrôle nerveux et hormonal. La muqueuse gastrique produit encore un GRP (Gastrin Releasing Peptide) qui stimule la production de gastrine.
Figure 5-19: A: Organisation d'un estomac de mammifère avec ses différents types de glandes. L'épithélium est limité vers le bas par une muqueuse musculaire et une sous-muqueuse en dessous de laquelle se trouvent 3 couches de muscles: obliques, circulaires et longitudinaux.
Dans la région cardiaque, on trouve essentiellement des cellules à mucus, comme dans la région œsophagienne squameuse. On y trouve en plus quelques cellules pariétales sécrétant de l'acide chlorhydrique. Dans la région gastrique, les cellules pariétales et principales sont abondantes. Ce sont en fait le corps de l'estomac et la région fundique qui participent le plus aux phénomènes de digestion. Dans la région pylorique, les glandes sont riches en cellules à mucus ainsi qu'en cellules endocrines. C'est dans cette région que sera essentiellement produite la gastrine, à partir de cellules dites "G".
4.1.3. Les mouvements de l'estomac
Ces mouvements n'ont été bien étudiés jusqu'à présent que chez l'homme et
quelques autres mammifères, le chien et quelques ruminants notamment. Nous nous
référerons donc essentiellement à ces travaux.
Du point de vue de la motricité, on distingue généralement deux parties dans l'estomac. Un premier tiers antérieur, l'estomac proximal, qui sert à recevoir la nourriture; les deux tiers restant constituent l'estomac distal où va s'effectuer l'essentiel du malaxage.
Les mouvements sont dus à trois couches de muscles lisses: circulaires, longitudinaux et obliques. Les contractions sont contrôlées par voie nerveuse réflexe et par voie hormonale (figures 5-20 et 5-21).
Figure 5-20: Les contractions gastriques et leur
contrôle.
Figure 5-21: Représentation schématique des voies nerveuses de contrôle de l'estomac chez les vertébrés. [Excitatrices: cholinergiques (––––– ); inhibitrices: adrénergiques ( ––––– ) et non adrénergiques ( ––––– )]. P: parasympathique, noyau moteur dorsal et nerf vague; S: sympathique, via ganglion céliaque; E: estomac. Chez les poissons et les amphibiens, les efférences parasympathiques vagales sont purement inhibitrices, les fibres cholinergiques sympathiques sont, elles, purement excitatrices. Chez les reptiles et les mammifères, il existe des efférences parasympathiques excitatrices et des efférences sympathiques devenant inhibitrices. D'après Stevens 1988, modifié.
L'estomac proximal sert avant tout à recevoir et à stocker la nourriture ingérée. C'est également à son niveau qu'est sécrété l'essentiel du suc gastrique. Il n'y a guère de mouvements musculaires importants dans cette région où le brassage du bol alimentaire est minime. Elle est seulement le siège de variations lentes de tonus qui interviennent dans la réception de la nourriture et dans son mouvement vers la partie distale. Au départ, la déglutition provoque l'ouverture réflexe du cardia et un relâchement des fibres musculaires par voie vagale. Ce réflexe de "relaxation réceptive" permet à l'estomac proximal de se distendre très largement au besoin. Le volume total chez l'homme peut ainsi passer de ± 50 ml au repos à 1500 ml après un repas copieux. La distension induit alors par voie sympathique les contractions lentes qui poussent le bol alimentaire vers l'estomac distal.
L'estomac distal a sa propre zone d'excitation. Il est le siège d'ondes péristaltiques puissantes qui vont effectuer, dans un mouvement de va-et-vient, un brassage important du bol alimentaire (chyme). Dans un premier temps, les ondes de contraction poussent le chyme vers le pylore. Cette région, très étroite ne contient qu'un faible volume (± 30 ml chez l'homme); elle agit comme un "filtre dynamique", ne laissant passer que des liquides et des petites particules par l'orifice pylorique qui s'ouvre légèrement sous la poussée du chyme. En général, chaque onde péristaltique n'éjecte que quelque ml de chyme vers l'intestin à travers le pylore. Le reste est reflué dans l'estomac par inversion du péristaltisme, les ondes de contractions repartent en sens inverse tout en fermant l'orifice pylorique.
L'intensité des ondes péristaltiques peut varier considérablement sous l'influence de facteurs nerveux et hormonaux mais leur fréquence paraît fixée par des cellules à activité rythmique spontanée (pacemakers) situées dans la couche longitudinale de muscles lisses. Le rythme est en général assez lent, de l'ordre de trois cycles par minute chez l'homme.
Les aliments déglutis sont ainsi brassés, mélangés au suc gastrique et réduits en particules d'un diamètre moyen de 0,3 mm qui passent dans l'intestin. Les temps de passage (transit) des aliments au niveau de l'estomac sont éminemment variables en fonction de leur nature chimique et de leur structure physique. Ces éléments, ainsi que le niveau de remplissage paraissent par ailleurs influencer l'importance de l'activité péristaltique. En général, chez l'homme, les temps de passage vont de 10 à 20 minutes pour les éléments liquides à 4 à 5 heures pour les aliments solides.
Chez les ruminants, on distingue un cycle de contractions reticulo-ruminal d'un cycle reticulo-omassal et d'un cycle abomassal. Les ondes partent du reticulum allant d'une part vers le rumen et d'autre part vers l'omassum pour revenir vers le reticulum, brassant ainsi le bol alimentaire et le faisant passer progressivement dans l'omassum. Périodiquement de fortes ondes péristaltiques parcourent l'omassum, poussent le bol alimentaire vers l'abomassum où il est brassé dans un mouvement de va-et-vient comme dans l'estomac des autres mammifères. Les ondes poussant le chyme de l'omassum vers l'abomassum ne se produisent pas au même rythme que les cycles. Elles sont par ailleurs stimulées par la distension de l'omassum et inhibées par celle de l'abomassum.
4.2.1. Organisation générale
Le dernier et généralement le plus long des segments du tube digestif, l'intestin
assure l'essentiel du processus de digestion par ses propres sécrétions ainsi que par
celles des deux glandes qui lui sont associées (pancréas et foie, voir 4.3
ci-après). Il assure également l'essentiel des processus d'absorption. Dans ce cadre, il
est la plupart du temps relativement long et présente des dispositifs augmentant encore
la surface absorbante (figure 5-22).
Les lamproies et la plupart des poissons, à l'exception des téléostéens évolués, ont un intestin assez gros et court dont la lumière est occupée par un repli longitudinal le plus souvent à disposition spiralée, ce qui ralentit considérablement le cheminement du bol alimentaire en augmentant son trajet. Cette structure, dite valvule spirale, peut atteindre l'axe central de l'intestin et se souder à elle-même en une columelle offrant l'aspect d'une vis. Cette disposition est particulièrement développée chez les requins où on peut compter jusqu'à 50 tours de spires (figure 5-22).
Figure 5-22: Les dispositifs d'augmentation de surface absorbante de l'intestin. A: Valvule spirale, ici chez un requin. B: Allongement considérable par plissement: plis de Kerckring (valvules conniventes), villosités et microvillosités. Chez l'homme, la surface est ainsi multipliée par 600.
Chez les tétrapodes et les téléostéens évolués, l'augmentation de surface est réalisée par allongement (figure 5-23). L'intestin fait ainsi de nombreuses circonvolutions et occupe l'essentiel de la cavité abdominale. On note également chez ces espèces, la présence de microvillosités, plissements de la face apicale des cellules épithéliales augmentant leur surface absorbante (figures 5-22 et 5-24). Chez les oiseaux et les mammifères, l'augmentation de surface est complétée par d'innombrables plissements et évaginations de la muqueuse de la portion grêle de l'intestin (figure 5-22). On aboutit ainsi à des surfaces épithéliales énormes, de l'ordre de 40 m2 chez l'homme pour une longueur de 7 m et un diamètre moyen de 3 cm par exemple.
Figure 5-23: Organisation générale de l'intestin
chez différentes espèces. LC: longueur du corps. A à C d'après Kardong 1995, modifié;
D à K d'après Stevens 1988, modifié.
Figure 5-24: A et B: L'épithélium de l'intestin
grêle. Schéma et microphotographie. - En A et B: villosités, entérocytes, cellules
caliciformes et glandes de Lieberkühn. Les produits de sécrétion des cellules
caliciformes apparaissent en clair au niveau de l'épithélium des villosités - En plus
sur le schéma glandes de Brunner spécifiques à la sous-muqueuse de la portion
duodénale et nodules lymphoïdes (cfr.
figure 5-25
). C et D: Vues en microscopie électronique, voir
figure 5-24: C et D (ci-après).
Figure 5-24: C et D: L'épithélium de l'intestin grêle. - En C vue en microscopie électronique d'une cellule caliciforme et de quelques entérocytes. - En D vue à fort grossissement de la zone apicale de deux entérocytes: d: desmosomes; f: microfilaments; L: lumière intestinale; mv: microvillosités; n: noyau; oz: zonula occludens (jonction étanche entre deux cellules).
L'intestin est divisé en régions distinctes morphologiquement et physiologiquement. Il s'ouvre sur l'estomac par le duodénum qui chez beaucoup de téléostéens est assorti de cæca pyloriques dont le nombre peut être très élevé (jusqu'à 1500, voir 4.1.1). Il se termine par une portion rectale qui débouche sur l'extérieur par un anus. Chez les amphibiens, les reptiles et les oiseaux, cette dernière partie reçoit les conduits des voies urinaires et génitales. Elle est alors organisée en un cloaque qui peut jouer un rôle non négligeable dans la récupération de l'eau et des ions provenant du système urinaire. Le cloaque n'existe pas chez les mammifères et n'est bien différencié chez les poissons que chez les sélaciens et les dipneustes. C'est à son niveau que s'ouvre la glande rectale jouant un rôle important dans l'excrétion de NaCl en milieu hyperosmotique (cfr. chapitre 1).
Chez les tétrapodes, l'intestin est divisé en intestin grêle et gros intestin ou colon. La division entre ces deux portions est marquée chez les oiseaux et les mammifères par la présence de cæca (2 chez les oiseaux, 1 chez les mammifères) dont le rôle dans la digestion est souvent mal défini. Chez les herbivores (autruche, lapin, cheval), leur développement peut devenir important. Ils participent à la digestion de la cellulose. Le temps de rétention du matériel végétal y est souvent assez long (cfr. 4.2.1).
4.2.2. Intestin grêle
L'intestin grêle est souvent plus long que le gros intestin. C'est à son niveau que
vont s'effectuer l'essentiel de la digestion et de l'absorption des nutriments, de l'eau
et des électrolytes.
Il commence par le duodénum, au niveau de l'extrémité inférieure de l'estomac; il se poursuit par le jéjunum et se termine
par l'iléon qui débouche dans le gros intestin. Ces divisions correspondent à des caractéristiques physiologiques et ultrastructurales des vertébrés supérieurs. Elles sont nettement moins nettes chez les vertébrés inférieurs.
4.2.3. Les sécrétions de la muqueuse
La surface épithéliale de l'intestin grêle est couverte de cellules absorbantes,
les entérocytes, dont la face apicale présente de très nombreuses évaginations, les
microvillosités, formant du côté lumière intestinale ce que l'on appelle la bordure en
brosse (figure 5-24 D). Ces microvillosités augmentent largement
la surface d'échange. Elles sont en plus capables de mouvements de rétraction (voir
chapitre 7), ce qui facilite le mouvement des molécules dans
l'épaisse couche muqueuse qui tapisse l'intestin. La fonction des entérocytes est
essentiellement absorbante. Ils jouent en plus un certain rôle digestif; comme nous
l'avons signalé précédemment; on trouve en effet différentes enzymes digestives
(dipeptidases, disaccharidases) sur la face apicale de ces cellules. On trouve également,
disséminées parmi les entérocytes, des cellules caliciformes (figure
5-24 C) qui sécrètent un abondant mucus qui va participer avec les sécrétions de
glandes spécialisées (voir plus loin) à la formation d'un gel muqueux (glycocalyx) qui
tapisse entièrement l'épithélium. Peu abondantes au niveau duodénal, elles augmentent
en nombre en allant vers l'iléon.
Chez les oiseaux et les mammifères, l'épithélium est largement pourvu de plis qui forment les villosités intestinales (figure 5-24 B). Chez les mammifères, la muqueuse et la sous-muqueuse sont encore plissées, essentiellement au niveau du jéjunum, très peu dans le duodénum et l'iléon, en de larges plis appelés plis ou valvules de Kerckring. Ces plis augmentent la surface d'absorption d'un facteur ± 1,3 par rapport à un tube lisse (figure 5-22). Chez l'homme, les villosités ont une forme aplaties dans le duodénum (figure 5-25) et prennent progressivement la forme de doigts de gant en allant vers l'iléon.
A la base des villosités s'ouvrent des cryptes communiquant avec la lumière des glandes dites de Lieberkühn dont le fond et les parois sont couverts de différents types cellulaires. Les parois présentent des cellules caliciformes sécrétant du mucus, des cellules indifférenciées d'où proviennent les nouvelles cellules villeuses et des cellules endocrines qui vont produire différentes hormones. Ces hormones, ainsi que d'autres produites par des neurones peptidergiques dont la distribution n'est pas toujours bien établie, vont agir sur les muqueuses gastrique et intestinale ainsi que sur les productions du foie et du pancréas. L'essentiel de ces substances est produit au niveau du duodénum et du jéjunum. L'effet de certaines d'entre elles a déjà été envisagé plus haut (voir 2.2 ci-avant). Il s'agit de la gastrine entérique qui stimule la production de suc gastrique mais surtout de la somatostatine, du GIP (Gastric Inhibitory Peptide) et de sécrétine qui inhibent la production de suc gastrique. Il s'agit par ailleurs de pancréozymine qui stimule avec la sécrétine les sécrétions pancréatiques et biliaires. L'intestin produit encore d'autres hormones tels le VIP (Vasoactive Intestinal Peptide) qui provoque une vasodilatation du système irriguant l'intestin, de la motiline et de la substance P qui stimulent les mouvements gastriques et intestinaux. L'iléon et le gros intestin sécrètent par ailleurs du PYY (Peptide Tyrosine, Tyrosine). Cette hormone, de découverte relativement récente (Tatemoto, PNAS 79, 2514, 1982) inhibe différents processus : mobilité intestinale, débit sanguin intestinal et pancréatique, sécrétions gastrique et pancréatique. Elle parait donc jouer un rôle important dans le retour aux conditions de repos après un repas. Elle serait sécrétée essentiellement en réponse à des stimuli nutritionnels et serait présente chez tous les vertébrés, des agnathes aux mammifères (Coulon, Amer. Zool. 35, 466, 1995).
Le fond des glandes de Lieberkühn est tapissé par des cellules dites de Paneth qui apparemment libèrent des enzymes digestives et des immunoglobulines. L'intestin grêle possède par ailleurs son tissu lymphatique immunocompétent propre; à savoir les plaques de Peyer qui sont particulièrement abondantes au niveau de l'iléon. Les plaques de Peyer apparaissent comme des dômes entre les villosités (figure 5-25). Elles sont recouvertes de cellules M (membraneuses) qui laissent passer les antigènes luminaux jusqu'aux follicules lymphoïdes sous-jacents qui sécrètent une IgA, ici protégée contre les enzymes digestives. Dans la portion duodénale de l'intestin, on trouve encore des glandes dites de Brünner. Elles sont localisées assez profondément dans la sous-muqueuse conjonctive et sécrètent un mucus riche en HCO3- qui neutralise l'acidité du chyme venant de l'estomac.
Figure 5-25: A: Les plaques de Peyer telles qu'elles apparaissent au microscope électronique à balayage après enlèvement de l'épithélium muqueux. B: Cellules membraneuses (M) couvrant un nodule lymphoïde (NL) en microscopie électronique à transmission.
On a décrit des glandes du type glande de Lieberkühn dans l'épithélium intestinal de la plupart des vertébrés, des batraciens aux mammifères. Il n'y en aurait pas chez les poissons, bien que l'on trouve dans l'intestin de ceux-ci l'essentiel des productions glandulaires décrites chez les autres espèces. Des structures de type glandulaire s'étendant dans la sous-muqueuse ont été décrites chez les gadidæ. Il semble cependant qu'elles ne contiennent pas de cellules différentes de celles de surface.
4.2.4. Gros intestin
Le gros intestin (colon) a pour fonction principale de compléter la réabsorption
d'eau et d'électrolyte entamée dans l'intestin grêle. Il concentre ainsi les résidus
alimentaires et aboutissant à la formation des fèces. Il est exempt de plis et de
villosités. La muqueuse est plus riche en cellules caliciformes que celle de l'intestin
grêle. Elle sécrète un gel muqueux abondant qui favorise le glissement des particules
alimentaires non digérées et des fèces. Il englobe également la flore de
micro-organismes présents en abondance à ce niveau. Les cellules absorbantes présentent
des microvillosités nettement plus courtes et moins nombreuses que dans l'intestin
grêle. Il y a aussi beaucoup moins de cellules endocrines. La sous-muqueuse est par
contre riche en nodules lymphoïdes. Dans la région anale, la muqueuse forme une série
de larges plis longitudinaux qui permettent en se distendant un stockage de la matière
fécale.
4.2.5. Les mouvements intestinaux
La muqueuse de l'intestin grêle est entourée de couches de muscles circulaires
(internes) et longitudinaux (externes). Ces muscles sont sous contrôle du parasympathique
qui les stimule et du sympathique qui provoque indirectement leur relâchement par
inhibition de fibres parasympathiques au niveau de groupes de neurones situés entre les
couches de muscles circulaires et longitudinaux (plexus nerveux myentérique ou
d'Auerbach). Il existe par ailleurs un plexus sous-muqueux qui contient les neurones
sensitifs des chémo-, mécano- et tensorécepteurs de la muqueuse.
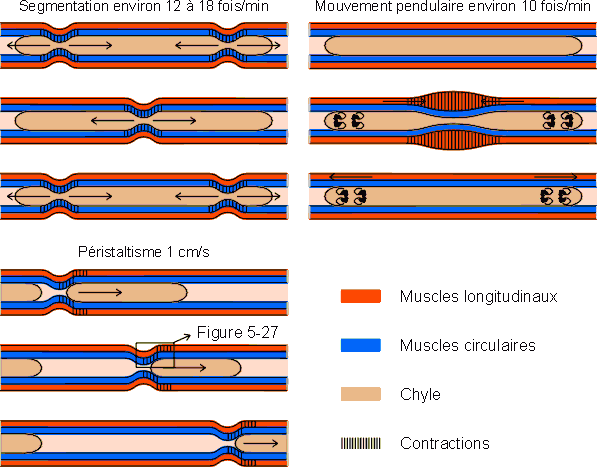
Outre les mouvements propres des microvillosités et des villosités permettant un contact étroit entre la muqueuse et les éléments du bol alimentaire, on peut distinguer trois types de mouvements servant à brasser et à faire avancer le chyle dans l'intestin grêle. On note ainsi des mouvements pendulaires mettant en jeu la musculature longitudinale, des mouvements de segmentation mettant en jeu les muscles circulaires et des mouvements péristaltiques poussant le chyle vers le gros intestin (figure 5-26). Le péristaltisme est déterminé par les tensorécepteurs qui déclenchent simultanément un resserrement de la lumière en amont du bol alimentaire et un élargissement en aval (figure 5-27). Le péristaltisme se marque essentiellement entre les périodes de prise d'aliments. Les mouvements de brassage priment largement au cours de l'alimentation et pendant la digestion.
Figure 5-26: Les mouvements de l'intestin grêle
chez l'homme.
Figure 5-27: Rôle des tensorécepteurs dans le péristaltisme intestinal des mammifères. Des fibres sensibles, partant des récepteurs, font relais sur des interneurones qui stimulent des nerfs moteurs activant les muscles longitudinaux et relâchant les muscles circulaires vers l'avant du bol alimentaire et, à l'inverse, inhibant les muscles longitudinaux et activant les muscles circulaires vers l'arrière. Les récepteurs sont excités par la distension que provoque le passage du bol.
Le gros intestin est le siège de deux types de mouvements : de fortes constrictions transversales brassant le bol alimentaire et un péristaltisme de masse se produisant uniquement à intervalles plus ou moins longs (de 2 à 4 heures chez l'homme). Dans ce mouvement, le colon se resserre brusquement autour de la partie la plus avancée du contenu intestinal et l'entraîne rapidement vers la partie terminale de l'intestin. Ces mouvements sont dus à un réflexe gastrocolique vagale et à des hormones intestinales.
Au fur et à mesure du remplissage du rectum supérieur, des tensorécepteurs sont stimulés qui déclenchent le besoin de déféquer. La défécation implique le relâchement des sphincters de l'anus qui sont sous commande réflexe; un contrôle conscient via des fibres motrices permet un retard temporaire.
La durée du transit intestinal de même que la fréquence des défécations sont éminemment variables en fonction des espèces. Le temps de passage de la nourriture peut ainsi varier de moins d'une heure chez de petites espèces (oiseaux, musaraignes) à plusieurs centaines d'heures (serpents, koala). Il est en général plus long chez les herbivores que chez les carnivores. Les temps de transit sont également fonction du type de nourriture, beaucoup plus rapide pour les fluides que pour les aliments particulaires ( figure 5-28A). Les temps de rétention dans les différentes parties de l'intestin sont également éminemment variables en fonction de l'espèce et du type de nourriture. Ils sont particulièrement longs dans les cæca chez les herbivores non ruminants. Ainsi chez le lapin par exemple, un repas de fibres marquées avec un radio-isotope se retrouvera essentiellement dans le cæcum intestinal dès après 2 heures. Après 24 h., plus de 60 % du marqueur seront toujours dans le cæcum alors que 20 % sont apparus dans les fèces, les 20 % restants étant distribués entre les autres parties de l'intestin (figure 5-28B). La longue rétention dans le cæcum permet la digestion du matériel végétal fibreux par les bactéries symbiotiques.
Chez la plupart des espèces, vertébrés comme invertébrés, le tube digestif est muni de glandes annexes qui jouent un rôle essentiel dans les processus digestifs. Elles sont toujours à l'étude chez les invertébrés où les connaissances actuelles restent très fragmentaires et permettent peu de généralisations. Elles sont mieux étudiées chez les vertébrés, où l'on distingue deux structures essentielles : le foie et le pancréas.
4.3.1. Le pancréas
Le pancréas déverse dans la partie haute de l'intestin un suc relativement aqueux
riche en HCO3- et en enzymes digestives; il contient
également du mucus. A côté de ses fonctions digestives, le pancréas exerce également
chez tous les vertébrés une fonction endocrine importante dont nous reparlerons (cfr.
chapitre 9). Des îlots de cellules (îlots dits de
Langerhans) sécrètent en effet de l'insuline et du glucagon, deux hormones impliquées
dans la régulation de la glycémie et du métabolisme du glucose.
Les cyclostomes et la plupart des actinoptérygiens ont un pancréas diffus, ne constituant pas un organe anatomiquement bien défini. Chez ces espèces, les cellules sécrétrices sont disséminées dans l'épithélium intestinal et le long des vaisseaux du mésentère, entre le foie et l'intestin. Elles s'infiltrent même dans le foie, le long des canaux biliaires. Chez les autres espèces, le pancréas se présente comme une volumineuse glande en grappe, composée de glandes acineuses. Ces glandes ont une structure voisine de celle des glandes salivaires. Elles sont constituées de cellules acineuses qui déversent leurs produits de sécrétion dans un canal intercalaire ( figure 5-29). Les cellules du canal se prolongent dans la lumière des acini en cellules dites centroacineuses, une caractéristique distinguant les acini pancréatiques des acini salivaires. Comme dans le cas des glandes salivaires, la sécrétion des cellules acineuses peut être modifiée par les cellules canalaires. Il y a généralement ajout de HCO3-, retrait d'eau et modification de concentration en Na+, K+ et Cl-. Les enzymes digestives produites par les cellules acineuses sont inactives et stockées dans de gros granules (grains de zymogène, figure 5-29B). Ces granules ont été très étudiés au cours des premiers travaux ultrastructuraux concernant la synthèse des protéines; le système pancréatique a en effet largement servi de modèle dans ces recherches.
Figure 5-29: Les acini du pancréas exocrine. A: Schéma. B: Vue en microscopie à transmission. CA: cellules acineuses avec granules de sécrétion (grains de zymogène, G); CC: partie de cellule centroacineuse bordant le canal intercalaire (CI).
4.3.2. Le foie
Le foie est souvent le viscère le plus volumineux des animaux. Il est essentiellement
constitué d'une masse de cellules épithéliales (parenchyme) traversée par des
capillaires veineux. Ces cellules sont disposées en un ensemble de lames ou travées
faites chez la plupart des espèces de deux épaisseurs de cellules ou d'une seule
(mammifères, quelques oiseaux - figure 5-30). Chez certains
mammifères (porc, ours, dromadaire), des cloisons conjonctives divisent le foie en
lobules centrés sur les extrémités des veines hépatiques. Le foie peut par ailleurs
être divisé en larges lobes; c'est le cas chez différents mammifères, comme l'homme
notamment.
Figure 5-30: Organisation schématique du tissu hépatique d'un foie lobulé de mammifère. En coupe, le lobule hépatique à l'aspect d'un hexagone dont les angles sont entourés d'une triade porte amenant le sang oxygéné par des branches de l'artère hépatique et les produits du métabolisme et de la digestion par des branches de la veine porte. Ce système est en connexion avec les lumières des capillaires sinusoïdes qui entourent les travées d'hépatocytes (beiges en B). Les produits du métabolisme hépatique se déversent dans les sinusoïdes et sont drainés par la veine centrolobulaire. La sécrétion biliaire est déversée dans de fins canalicules entre les hépatocytes et drainée par les canalicules biliaires dans les canaux biliaires des triades porte.
Les voies biliaires débutent dans le parenchyme hépatique par des canalicules sans parois propres; il s'agit essentiellement de l'espace intercellulaire qui forme un réseau en relation avec des canaux biliaires intra-hépatiques qui se réunissent en canaux (extra) hépatique aboutissant à un canal cholédoque qui conduit la bile jusqu'au duodénum (voir 2.4).
Le foie exerce une quantité de fonctions dont certaines ayant trait au métabolisme, à la détoxification ou à l'excrétion de différents composés pour l'ensemble de l'organisme. Il joue par ailleurs un rôle essentiel dans l'assimilation des lipides et la gestion des réserves de glycogène et de graisses. Dans ces cadres, il trie et gère l'essentiel des produits entrant dans l'organisme d'une part et provenant du métabolisme cellulaire d'autre part. L'importance physiologique que revêt ainsi le foie s'explique par ses remarquables potentialités biochimiques liées à un exceptionnel équipement enzymatique ainsi qu'à une situation privilégiée sur l'appareil circulatoire. Nous reviendrons sur certains rôles du foie dans d'autres chapitres. Les capillaires sinusoïdes du foie contiennent par ailleurs de très nombreux macrophages (cellules de Kupffer des histologistes) qui jouent un rôle essentiel dans la phagocytose et l'élimination des vieux globules rouges (voir chapitre 6) mais aussi de particules virales, de bactéries ou de complexe immuns.
Dans sa fonction digestive, le foie produit la bile qui joue un rôle important dans la digestion des lipides en favorisant leur émulsion (voir 2.4 ci-avant). Comme nous le verrons au chapitre suivant, la bile sert aussi à l'élimination via l'intestin de certains produits détoxifiés par le foie. La plupart des vertébrés ont une vésicule biliaire pouvant stocker et concentrer la bile. Ce n'est cependant pas le cas des lamproies, chez lesquelles elle disparaît à la métamorphose, de certains téléostéens, reptiles, oiseaux et mammifères tel le cheval, le rat, le phoque ou le cerf (Stevens, 1988).