2.2.1. Enzymes majeures impliquťes
2.2.2. Modalitťs et contrŰle de sťcrťtion
2.2.3. Aspects comparťs
2.3.1. Enzymes impliquťes
2.3.2. Modalitťs et contrŰle de sťcrťtion
2.3.3. Aspects comparťs
a. glycogŤne et amidon
b. cellulose
c. chitine
d. autres polysaccharides
2.4.1. Enzymes impliquťes
2.4.2. Modalitťs et contrŰle de sťcrťtion
2.4.3. Aspects comparťs
La digestion concerne l'ensemble des processus au cours desquels les aliments ingťrťs, constituťs en large part d'ťlťments non directement utilisables par les cellules, sont rťduits par hydrolyse enzymatique en nutriments utilisables. La digestion peut Ítre extracellulaire, intracellulaire ou de contact.
Le systŤme de digestion intracellulaire implique l'incorporation d'ťlťments ingťrťs par phagocytose aux vťsicules lysosomiales dans lesquels ils seront hydrolysťs. L'ensemble du processus est encore loin d'Ítre clair et relŤve essentiellement de la physiologie cellulaire (cfr. livre 2). Le systŤme de digestion extracellulaire implique une sťcrťtion d'enzymes digestives dans un espace dťterminť, extťrieur aux cellules, oý les ťlťments ingťrťs sont hydrolysťs complŤtement ou partiellement. Si la digestion intracellulaire reste essentielle dans le cadre des relations entre cellules au sein d'un organisme, elle n'a d'importance dans la digestion d'ťlťments extťrieurs au niveau organismique que chez quelques groupes primitifs (cœlentťrťs, plathelminthes). Elle est de peu d'importance, voire inexistante dans tous les autres groupes. Certains ťlťments ne sont que partiellement dťgradťs par le systŤme extracellulaire. Dans ces cas, les molťcules rťsultantes sont souvent digťrťes par des enzymes associťs aux membranes plasmiques avant d'Ítre assimilťes. Certains parlent dans ce cas de digestion de contact.
La digestion extracellulaire s'effectue dans certains endroits du tractus digestif ( figure 5-1). L'intestin intervient dans tous les cas, l'estomac dans de nombreux cas et la cavitť buccale dans quelques cas seulement.
La digestion met en jeu des enzymes et des structures diffťrentes selon la nature chimique des aliments ingťrťs: protťines, lipides ou hydrates de carbone (glucides). La plupart des ťtudes concernant les mťcanismes impliquťs sont relatives aux mammifŤres. Notre examen des systŤmes digestifs concernera donc essentiellement ce groupe. Nous nous rťfťrerons autant que faire se peut aux donnťes obtenues chez d'autres espŤces: elles sont malheureusement souvent peu nombreuses, parfois rares voire inexistantes, surtout pour les invertťbrťs.
2.2.1. Enzymes majeures impliquťes
Les protťines sont formťes de chaÓnes polypeptidiques plus ou moins longues,
elles-mÍmes constituťes d'acides aminťs associťs entre eux par une liaison amide
substituťe ou liaison peptidique. Cette liaison implique le groupement NH2
d'un acide aminť et le groupement COOH d'un autre.
La digestion des protťines va donc essentiellement consister en l'hydrolyse de ces liaisons par des enzymes digestives appelťes peptidases (figure 5-8). Le rťsultat final est la formation d'acides aminťs et de dipeptides qui seront absorbťs au niveau de l'intestin.
|
R
R
NH2 | | | HOOC - CH - NH - C - CH - NH2 +H2O ģ 2R - CH - COOH || O |
Les peptidases sont classťes en endo et exopeptidases selon qu'elles attaquent une liaison peptidique ŗ l'intťrieur ou ŗ l'extťrieur d'une protťine. Les exopeptidases sont elles-mÍmes classťes en carboxypeptidases et aminopeptidases selon que le groupement terminal du dernier acide aminť de la chaÓne ŗ cliver est acide (groupement COOH libre) ou aminť (groupement NH2 libre).
Les carboxypeptidases sont sťcrťtťes par le pancrťas sous forme inactive : les procarboxypeptidases. Ces procarboxypeptidases sont activťes dans le suc pancrťatique sous l'action d'une autre enzyme digestive, la trypsine; elle aussi sťcrťtťe au niveau pancrťatique. Les aminopeptidases sont sťcrťtťes par les cellules intestinales directement sous forme active.
Les endopeptidases sont au nombre de trois : pepsine, trypsine et chymotrypsine ( figure 5-8).
Les pepsines agissent sur les liaisons peptidiques dans lesquelles le groupement aminť provient d'un acide aminť aromatique (tyrosine ou phťnylalanine) et le groupement acide d'un acide aminť dicarboxylique (aspartate ou glutamate). Elles sont sťcrťtťes au niveau de l'estomac sous formes inactives, les pepsinogŤnes, par des cellules dites principales ou ŗ zymogŤne (voir plus loin : 4.2). Les pepsinogŤnes sont activťs en pepsines par le pH gastrique acide, lorsqu'il est compris entre 2 et 4.
La trypsine agit sur les liaisons peptidiques dont le groupement carboxylique provient de la lysine ou de l'argine alors que la chymotrypsine clive avec le plus d'efficacitť les liaisons peptidiques dont la fonction carboxylique est fournie par les acides aminťs aromatiques (tyrosine ou phťnylalanine). Les deux enzymes sont sťcrťtťes sous forme inactive (trypsinogŤne et chymotrypsinogŤne) par le pancrťas. Le chymotrypsinogŤne est activť par la trypsine et le trypsinogŤne par une peptidase spťcifique sťcrťtťe par la muqueuse intestinale : l'entťrokinase.
Figure 5-8: Les enzymes de la digestion des protťines. Explications dans le texte. * et (Glycine) : la nature de l'acide aminť intervenant ici n'est pas importante. Les liaisons peptidiques apparaissent en bleu.
2.2.2. Modalitťs et contrŰle de sťcrťtion
La digestion des protťines; de mÍme que celle des autres aliments est contrŰlťe
dans le cadre d'une sťquence gťnťrale impliquant diffťrents facteurs. Ces ťlťments
sont considťrťs essentiellement dans la section 4.1.2 et
4.2.2 ci-aprŤs. BriŤvement, l'hydrolyse des protťines
commence au niveau de l'estomac avec la production d'un suc gastrique acide contenant les
pepsinogŤnes que l'aciditť ambiante active en pepsines. Cette sťcrťtion est
influencťe par des facteurs :
a) nerveux : des effťrences vagales activent la production de HCl et de pepsinogŤnes mais aussi de gastrine, une substance qui va elle-mÍme stimuler la production de suc gastrique. Le rťflexe vagale est essentiellement en rapport avec des stimuli gustatifs, olfactifs ou visuels associťs ŗ la prise de nourriture. Il peut cependant faire facilement l'objet d'un conditionnement (voir systŤme nerveux) permettant ainsi de nombreuses autres influences. L'ťtat psychique, le stress, la peur peuvent ťgalement influencer la sťcrťtion du suc gastrique.
b) locaux : le contact de certains ťlťments du bol alimentaire (acides aminťs notamment) dans les parties profondes de l'estomac provoque la sťcrťtion de gastrine par la muqueuse pylorique. Il en va de mÍme pour la distension de la partie supťrieure de l'estomac qui a lieu lors de l'arrivťe du bol alimentaire. La gastrine va activer par voie sanguine la production d'acide chlorhydrique par des cellules se trouvant plus haut, dans l'estomac dit proximal (cfr. 4.1.2). Un pH trŤs acide inhibera la production de gastrine en rťtroaction, alors qu'un pH insuffisamment acide l'activera.
c) hormonaux : diffťrentes substances produites par les muqueuses gastriques ou intestinales et sťcrťtťes dans le flot sanguin vont agir sur la production de suc gastrique. C'est le cas notamment de la gastrine, de l'histamine ou du GRP (Gastric Releasing Peptide), produits par la muqueuse gastrique et qui sont activateurs. C'est aussi le cas de la sťcrťtine et du GIP (Gastric Inhibitory Peptide), deux substances produites par la muqueuse de la partie haute (duodťnale) de l'intestin au contact de certains facteurs du chyme provenant de l'estomac: pH acide, prťsence de glucose, acides aminťs ou lipides. Ces substances inhibent la production de suc gastrique.
Une premiŤre attaque des protťines par les pepsines a donc lieu au niveau gastrique en milieu acide. Le reste de la sťquence a lieu au niveau de l'intestin oý vont intervenir trypsine, chymotrypsine, amino et carboxypeptidases. Ces enzymes contrairement aux pepsines, sont actives ŗ des pHs voisins de la neutralitť. Le bol alimentaire sera donc ramenť ŗ pH neutre ŗ l'entrťe de l'intestin par des sťcrťtions de HCO3 - venant du pancrťas et des glandes duodťnales (cfr. 4.2.2 et 4.3.1 ci-aprŤs).
L'arrivťe du bol alimentaire de l'estomac (chyme) dans le duodťnum va induire la production par la muqueuse intestinale : a) vers le systŤme circulatoire, de diffťrentes hormones notamment la sťcrťtine et la pancrťozymine. Ces deux hormones ainsi que les stimulations vagales, vont induire la production du suc pancrťatique contenant notamment des procarboxypeptidases, des chymotrypsinogŤnes et des trypsinogŤnes. La sťcrťtine inhibe par ailleurs la sťcrťtion gastrique de HCl. b) vers la lumiŤre intestinale, de l'entťrokinase, une enzyme qui activera le trypsinogŤne en trypsine. La trypsine activera ŗ son tour le chymotrypsinogŤne en chymotrypsine et les procarboxypeptidases en carboxypeptidases. Les enzymes protťolytiques pancrťatiques deviennent donc actives dans l'intestin. Elles fonctionnent dans un milieu ŗ pH neutre. Dans ce cadre, la sťcrťtion pancrťatique contient du HCO3- qui la rend basique et qui a pour effet de tamponner le bol alimentaire dans l'intestin ŗ des pHs proches de la neutralitť. c) La production des aminopeptidases par la muqueuse intestinale.
L'ensemble de cette sťquence va rťsulter en l'hydrolyse de l'essentiel des protťines en acides aminťs, di- et tripeptides. Ces peptides seront clivťs en acides aminťs par des peptidases fixťes ŗ la membrane plasmique de cellules intestinales, essentiellement de la portion grÍle. L'absorption des acides aminťs a lieu dans l'intestin et fait intervenir des cotransporteurs Na-acides aminťs (cfr. 5.1 et surtout physiologie cellulaire).
Bon nombre d'aliments d'origine carnťe contiennent de l'ťlastine, une protťine fibreuse ťlastique prťsente notamment dans les tissus conjonctifs, les ligaments et les parois vasculaires. Cette protťine est mal digťrťe par les protťases classiques. La sťcrťtion pancrťatique des mammifŤres contient une ťlastase spťcifique. Elle est produite sous forme de proťlastase inactive et est activťe par la trypsine.
2.2.3. Aspects comparťs
Les recherches concernant les enzymes protťolytiques dans les groupes autres que
mammaliens ne sont que trŤs partielles. En ce qui concerne les vertťbrťs, des enzymes
ŗ activitť de type pepsine, trypsine et chymotrypsine ont ťtť mises en ťvidence chez
les oiseaux, diffťrents reptiles (tortues, lťzards) ainsi que quelques amphibiens,
anoures et urodŤles. Chez les anoures, la pepsine stomacale n'apparaÓt qu'ŗ la
mťtamorphose. Elle est absente chez le tÍtard.
Chez les poissons, on trouve les diffťrentes enzymes protťolytiques pancrťatiques et intestinaux. La prťsence d'enzymes de type pepsine est variable. Elles n'existent pas chez les poissons dťpourvus d'estomac (sťlaciens, chimŤres, dipneustes). Chez les tťlťostťens, la pepsine n'est pas toujours prťsente, mÍme chez les espŤces possťdant un estomac bien diffťrenciť sťcrťtant de l'acide chlorhydrique. Chez les espŤces qui la possŤdent, il semble que les cśca pyloriques soient un site important de production.
Les enzymes protťolytiques ont ťtť hautement purifiťes chez quelques espŤces et leur sťquence en acides aminťs est connue. Il existe, semble-t-il un haut degrť d'homologie entre les diffťrentes peptidases des diffťrents vertťbrťs.
En ce qui concerne les invertťbrťs, la situation est moins claire, avec des rťsultats souvent fragmentaires et prťliminaires. Des activitťs de type trypsine et/ou chymotrypsine ont ťtť mises en ťvidence dans tous les groupes jusqu'aux urocordťs ŗ l'exception des hirudinťes (sangsues). La prťsence d'exopeptidases a ťgalement ťtť dťmontrťe chez de nombreuses espŤces. Dans la plupart des cas, la source prťcise de ces enzymes est inconnue.
La prťsence d'enzymes de type pepsine est douteuse, mÍme chez les espŤces, comme les crustacťs dťcapodes par exemple, possťdant un estomac bien diffťrenciť. En gťnťral, le pH dans le tractus digestif des invertťbrťs varie entre 5 et 8 ce qui exclut la prťsence d'enzymes travaillant ŗ pH trŤs bas.
L'absence totale d'endopeptidases chez les sangsues alors que leurs aliments essentiels sont protťiques (protťines du plasma et des globules rouges sanguins) est curieuse. Elle pourrait rendre compte de la lenteur avec laquelle les repas de sang sont digťrťs chez ces espŤces. Elles ne disposeraient en fait que d'exopeptidases aidťes plus ou moins efficacement par des enzymes d'origine bactťrienne. Chez les insectes suceurs de sang, on trouve par contre un arsenal complet (sauf pepsine) de peptidases ainsi qu'une protťinase particuliŤre ayant une activitť particuliŤrement ťlevťe contre l'hťmoglobine.
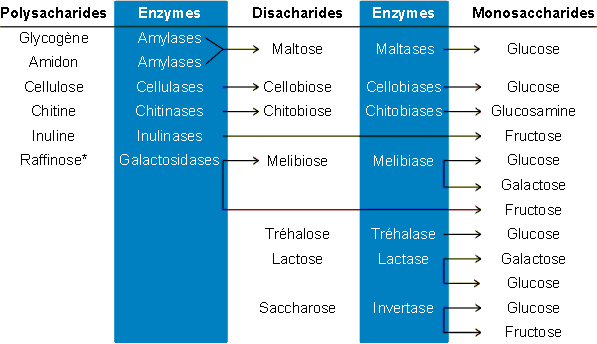
2.3.1. Enzymes impliquťes
Les enzymes hydrolytiques impliquťes dans la digestion des sucres sont assez
spťcifiques: elles seront donc plus nombreuses que dans le cas des protťines ou des
lipides oý il existe des enzymes "gťnťrales" actives sur un type de liaison
plutŰt que sur un composť donnť. On distingue des enzymes hydrolysant les
polysaccharides (polyholosides) d'autres attaquant les disaccharides et trisaccharides (di
et triholosides). En ce qui concerne les polysaccharides, le glycogŤne et l'amidon sont
digťrťs par des amylases, la cellulose par des cellulases, l'inuline par des inulinases,
la chitine par des chitinases, etc. En ce qui concerne les dissacharides, le maltose est
clivť par des maltases, le cellobiose par des cellobiases, le lactose par des lactases,
etc. Le raffinose est le plus rťpandu des trisaccharides et pratiquement le seul a avoir
ťtť ťtudiť. Il est hydrolysť par des galactosidases (figure 5-9).
Figure 5-9: Les enzymes de la digestion des glucides. * Le raffinose est un trisaccharide.
L'arsenal enzymatique existant chez les animaux pour digťrer les glucides est en gťnťral trŤs incomplet. Bon nombre d'enzymes intervenant ici seront donc d'origine exogŤne, provenant de bactťries et/ou de protozoaires logťs dans l'estomac ou l'intestin. La digestion de la cellulose par exemple repose essentiellement sur des enzymes d'origine exogŤne. Elle sera envisagťe plus loin dans le cadre des apports symbiotiques de nutriments (cfr. section 3). Les diffťrentes polysaccharidases endogŤnes sont essentiellement sťcrťtťes par le pancrťas. Certaines espŤces peuvent ťgalement avoir des polysaccharidases salivaires. Ainsi la salive humaine contient une a amylase (ptyaline). Les disaccharidases endogŤnes paraissent localisťes sur la muqueuse de l'intestin, dans sa portion grÍle essentiellement. Les enzymes chitinolytiques, chitinase et chitobiase, ont ťtť mises en ťvidence chez diffťrentes espŤces de mammifŤres. Les chitinases sont gastriques et/ou pancrťatiques alors que les chitobiases seraient plutŰt intestinales, parfois gastrique ťgalement. Les activitťs chitinolytiques sont gťnťralement trŤs ťlevťes chez les insectivores, elles sont nulles chez l'homme, le lapin ou le mouton. Une hyaluronidase a ťtť mise en ťvidence dans la salive de tous les mammifŤres ťtudiťs, l'activitť de l'enzyme ťtant plus ťlevťe chez les carnivores que chez les herbivores.
2.3.2. Modalitťs et contrŰle de sťcrťtion
La sťcrťtion des polysaccharidases salivaires et pancrťatiques fait intervenir
des facteurs nerveux, hormonaux et locaux (voir plus haut et figure 5-13)
Les influences nerveuses sont vagales (parasympathiques) pour le pancrťas, sympathiques
et parasympathiques pour les glandes salivaires. La production de suc pancrťatique est en
fait essentiellement contrŰlťe par les hormones sťcrťtine et pancrťozymine produites
par la portion haute de l'intestin au passage du bol alimentaire acide provenant de
l'estomac (figure 5-13).
Les oses rťsultant de l'activitť hydrolytique sont absorbťs au niveau intestinal essentiellement par des cotransporteurs Na-oses, pour le glucose et le galactose en tout cas. Il semble que le passage du fructose soit essentiellement diffusionnel (cfr. 5.1).
2.3.3. Aspects comparťs
a. glycogŤne et amidon
Ces composťs sont des produits de rťserve majeurs des animaux (glycogŤne) et des
plantes supťrieures (amidon) Ils sont tous deux constituťs d'unitťs glucose en liaison
a -1,4 pour les chaÓnes droite et a -1,6
au niveau des points de branchement pour les chaÓnes ramifiťes. Dans l'amidon, les
chaÓnes droites portent le nom d'amylose et les chaÓnes ramifiťes d'amylopectine.
GlycogŤne et amidon sont hydrolysťs en maltose par des amylases puis en glucose par des disaccharidases spťcifiques. On a trouvť des amylases pancrťatiques et des maltases intestinales chez tous les vertťbrťs ťtudiťs jusqu'ŗ prťsent, des sťlaciens aux mammifŤres. On a ťgalement trouvť ces enzymes dans le tube digestif de tous les invertťbrťs testťs sauf les sangsues.
b. cellulose
La cellulose est un polysaccharide de structure majeur des vťgťtaux. C'est un
constituant essentiel des parois cellulaires. Constituťe de longues chaÓnes d'unitťs
glucose en liaison b -1,4, elle n'est pas hydrolysable par les
amylases du tube digestif des vertťbrťs. La cellulose est digťrťe chez eux par des
enzymes d'origine exogŤne. La situation est moins claire chez les invertťbrťs oý l'on
trouve des cellulases endogŤnes ŗ activitť rťduite chez certaines espŤces dans la
plupart des groupes. Dans la plupart des cas, ces activitťs ne sont pas suffisantes pour
rendre compte d'une digestion effective et il existe une contribution d'origine exogŤne
importante.
La contribution exogŤne ŗ la digestion de la cellulose est dans la plupart des cas le fait de bactťries et d'unicellulaires prťsents en symbiotes dans diffťrentes parties du tractus digestif; cśca œsophagiens, estomac, intestin et/ou cśca intestinaux (voir section 3 ci-aprŤs). Certaines espŤces peuvent obtenir une aide non nťgligeable d'organismes se trouvant normalement dans leur alimentation et prťsentant une activitť cellulolytique ťlevťe. Chez les poissons, il pourra s'agir d'invertťbrťs prťsents sur les plantes par exemple. Dans d'autres cas, il peut s'agir de champignons. Ainsi, l'isopode marin xylophage Limnoria tripunctata se nourrit gťnťralement de vieux bois infestťs de champignons eux-mÍmes xylophages. Son tube digestif ne prťsente pas de flore microbienne particuliŤre. De mÍme, certaines espŤces de termites comme Macrotermes natalensis trouvent une aide ŗ la digestion de la cellulose dans les champignons qu'elles cultivent (Termitomyces sp.).
c. chitine
La chitine est un polymŤre de N - acťtylglucosamine dont les molťcules sont unies
en enchaÓnements linťaires par des liaisons b -1,4. Associťe
ŗ des sels minťraux et ŗ des protťines, elle constitue l'exosquelette des arthropodes.
Des chitinases et des chitobiases ont ťtť mises en ťvidence dans tous les groupes de vertťbrťs et dans de nombreux groupes d'invertťbrťs. L'activitť chitinolytique est en gťnťral ťlevťe chez les insectivores ou chez des espŤces mangeant occasionnellement insectes ou crustacťs. Elle est basse ou nulle chez les autres, notamment chez les herbivores. Chez les vertťbrťs, les chitinases sont essentiellement d'origine gastrique et/ou pancrťatique. Dans bien des cas, comme pour la cellulose, il semble que la dťcomposition par enzymes exogŤnes puisse contribuer de maniŤre non nťgligeable ŗ la digestion de la chitine.
d. autres polysaccharides
Les recherches concernant d'autres composťs sont restťes peu nombreuses ŗ ce jour.
Des hyaluronidases ont ťtť mises en ťvidence chez les poissons, dans l'estomac chez certains, dans l'intestin chez d'autres. La sangsue Hirudo medicinalis possŤde une hyaluronidase salivaire trŤs active qu'elle injecte, en mÍme temps qu'un anticoagulant, dans la blessure qu'elle fait aux animaux qu'elle attaque. L'acide hyaluronique est un constituant important des conjonctifs. C'est un polymŤre linťaire de disaccharides en liaison b -1,4 dont chaque unitť est constituťe d'une acide glucuronique et d'une N - acťtylglucosamine en liaison b -1,3. L'enzyme d'Hirudo hydrolyse uniquement ces derniŤres liaisons. Il diffŤre en cela des hyaluronidases des venins de serpents ou d'abeilles qui s'attaquent aux liaisons b -1,4 entre les disaccharides. La laminarine (glucoses en b -1,3) et l'acide alginique (chaÓnes d'acides mannuronique et guluronique) sont des constituants majeurs des algues brunes. Ces composťs ne paraissent pas pouvoir Ítre digťrťs par les vertťbrťs, y compris les poissons vivants parmi ces algues. Une activitť d'hydrolyse de la laminarine a pu Ítre mise en ťvidence dans le tube digestif de diffťrents invertťbrťs : anťmone de mer, nombreux polychŤtes, oligochŤtes, crustacťs, mollusques et ťchinodermes. De mÍme, des alginases sont prťsentes chez de nombreuses espŤces, essentiellement marines. Le niveau d'activitť des alginases comme des laminarinases paraÓt essentiellement fonction du rťgime alimentaire. Ces enzymes seraient d'origine endogŤne, quoique le lieu de production reste non prťcisť dans la plupart des cas.
2.4.1. Enzymes impliquťes
Les enzymes lipolytiques sont classťes en lipases et estťrases. Les lipases
hydrolysent les triglycťrides alors que les autres composťs lipidiques sont hydrolysťs
par des estťrases plus ou moins spťcifiques. Les triglycťrides (tri-acylglycťrols)
sont des constituants de rťserve essentiels chez les animaux. Ils reprťsentent en
gťnťral plus de 90 % des lipides du bol alimentaire chez les mammifŤres. Le reste est
constituť de phospholipides, cholestťrol, esters de cholestťrol et cťrides.
L'essentiel des enzymes lipolytiques provient du pancrťas oý elles paraissent sťcrťtťes sous forme de pro-enzymes inactives, activťes dans l'intestin par la trypsine. Chez diffťrentes espŤces, il existe ťgalement une lipase salivaire, produite au niveau des glandes sublinguales. Cette enzyme peut Ítre assez active. On estime ainsi que chez l'homme 20 ŗ 30 % des triglycťrides peuvent Ítre hydrolysťs par la lipase salivaire avant d'atteindre l'intestin. On trouve par ailleurs une lipase non spťcifique dans le lait maternel, ce qui apporte au nourrisson en allaitement la graisse du lait en mÍme temps que son enzyme de digestion. Cette enzyme peut hydrolyser les triglycťrides mais aussi les phospholipides, les esters de cholestťrol ainsi que de nombreux autres esters. Une lipase de mÍme type est ťgalement sťcrťtťe par le pancrťas en mÍme temps que les autres enzymes lipolytiques plus spťcifiques. Parmi ces derniŤres on trouve notamment une lipase 1 hydrolysant les triglycťrides au niveau des acides gras extťrieurs (1 et 3). L'acide gras restant, en position 2, est libťrť du glycťrol par une lipase 2 ou par la lipase non spťcifique (figure 5-10). Le pancrťas sťcrŤte encore une colipase dont le rŰle est de lever l'inhibition qu'exerce les sels biliaires sur l'activitť des lipases (voir ci-aprŤs).
Figure 5-10: Lipases 1 et 2 dans la digestion des triglycťrides.
2.4.2. Modalitťs et contrŰle de sťcrťtion
La sťcrťtion des enzymes lipolytiques salivaires et pancrťatiques fera intervenir
les facteurs nerveux, locaux et hormonaux que nous avons dťjŗ mentionnťs en envisageant
la digestion des glucides et des protťines (voir plus haut et figure
5-13).
Les lipides et leurs produits de dťgradation sont peu solubles dans l'eau alors que les enzymes lipolytiques sont hydrosolubles ce qui rend les contacts difficiles. De plus, dans les solutions aqueuses, les lipides s'agglomŤrent pour former de gros agrťgats diminuant largement le rendement de l'hydrolyse enzymatique puisque seules les molťcules situťes ŗ la surface des agrťgats sont accessibles aux enzymes. La digestion des lipides est rendue plus efficace par un processus d'ťmulsification transformant les agrťgats lipidiques en micelles de faibles diamŤtres (3 ŗ 6 nm seulement). Ce processus commence dans l'estomac avec le brassage mťcanique intense que subit le bol alimentaire. Il est complťtť dans l'intestin par un brassage mťcanique auquel s'ajoute l'action des sels biliaires. Ces sels vont en fait stabiliser l'ťmulsion produite par le brassage mťcanique. Leurs molťcules possŤdent des rťgions polaires et non polaires et adhťrent par ces derniŤres aux molťcules lipidiques. Les parties polaires, dirigťes vers la phase aqueuse ťtablissent des attractions avec l'eau et se repoussent ce qui maintient la division physique des micelles et forme une ťmulsion stable (figure 5-11). Chez les mammifŤres, les sels biliaires sont essentiellement des tauro et des glyco-conjuguťs d'acides biliaires. Ils sont synthťtisťs par le foie et stocker dans la vťsicule biliaire pour Ítre dťversťs dans le duodťnum lors du passage du bol alimentaire (voir aussi 4.3.2 et 5.3).
La production de sels biliaires et de bile par le foie est continue ( Ī700 ml/24h chez l'homme). La bile n'est cependant dťversťe dans le duodťnum que par intervalles, essentiellement lors du passage du bol alimentaire. La bile produite par le foie est en fait stockťe dans la vťsicule biliaire au niveau de laquelle elle est ťgalement concentrťe de 5 ŗ 10 fois par pompage d'eau. La vťsicule biliaire et le foie sont en relation avec la portion haute de l'intestin par le canal cholťdoque, fermť par un muscle constricteur (sphincter d'Oddi). La sťcrťtion biliaire est contrŰlťe par voie nerveuse rťflexe (vagale) mais surtout par voie humorale. La pancrťozymine libťrťe dans le sang par les cellules duodťnales lors du passage du bol alimentaire provoque en effet la contraction de la vťsicule biliaire et le rel‚chement du sphincter d'Oddi (effets cholagogues). La sťcrťtine a par ailleurs des effets stimulants sur la production de bile par le foie (effets cholťrťtiques).
Les sels biliaires dťversťs dans la portion antťrieure de l'intestin grÍle sont rťabsorbťs dans sa portion postťrieure (ilťon) aprŤs absorption des micelles lipidiques qu'ils ont aidťs ŗ stabiliser. Ils peuvent ainsi Ítre rťutilisťs ŗ la formation de bile en un cycle dit entťro-hťpatique (voir section 5.3).
Les sels biliaires sont synthťtisťs dans le foie ŗ partir d'acides biliaires dits primaires (acides cholique et dťsoxycholique) eux-mÍmes synthťtisťs dans le foie ŗ partir du noyau cyclopentanophťnanthrťnique dťrivť du cholestťrol (figure 5-11). Une partie de ces acides peut Ítre transformťe en acides biliaires secondaires dans l'intestin par action bactťrienne. Ces derniers, aprŤs rťabsorption participent dans le foie avec les acides biliaires primaires ŗ la formation de conjuguťs notamment avec de la glycine, de la taurine mais aussi des sulfates. Les conjuguťs sont salifiťs par des ions monovalents (Na et K essentiellement) pour former les sels biliaires.
Figure 5-11: Sels biliaires et formation de micelles lipidiques - Le noyau cyclopentanophťnanthrťnique et le cholestťrol, base des acides et sels biliaires - Les acides glyco et taurodťsoxycholique ont la mÍme structure que les conjuguťs de l'acide cholique prťsentťs ŗ ceci prŤs qu'en position 7 (colorťe), il y a un H au lieu d'un OH.
2.4.3. Aspects comparťs
On trouve des lipases et des estťrases chez tous les vertťbrťs. Elles sont
essentiellement d'origine pancrťatique. Chez certaines espŤces, on trouve ťgalement des
lipases gastriques et salivaires. Chez les serpents, les sťcrťtions des glandes ŗ venin
peuvent ainsi contenir des phospholipases et des cholinestťrases inhibant la transmission
nerveuse. L'origine des lipases actives dans le suc gastrique n'est pas claire. Il
n'existe en effet aucune dťmonstration probante ŗ l'heure actuelle des possibilitťs de
synthŤse de telles enzymes par la muqueuse gastrique. Peut Ítre s'agit-il de lipases
salivaires ou d'enzymes exogŤnes, amenťes avec le bol alimentaire ou produites par des
bactťries de l'estomac. Dans de nombreux cas, des enzymes d'origine bactťrienne
contribuent de faÁon non nťgligeable et parfois essentielle ŗ la digestion des lipides
du bol alimentaire (voir section 3).
Les vertťbrťs terrestres trouvent surtout des triglycťrides dans leur nourriture. Pour certaines espŤces marines, le zooplancton constitue une source essentielle de nourriture ou un appoint non nťgligeable. Ces animaux sont capables de digťrer les cires (esters d'alcools supťrieurs et d'acides gras non saturťs ŗ chaÓnes longues) se trouvant en abondance dans les animaux planctoniques. On ignore encore si les enzymes impliquťes sont essentiellement d'origine endogŤne ou exogŤne et s'il s'agit d'estťrases spťcifiques ou non spťcifiques.
Des lipases et des estťrases ont ťtť mises en ťvidence dans le tube digestif de la plupart des invertťbrťs ťtudiťs jusqu'ŗ prťsent. Le suc digestif du priapulide Priapulus caudatus paraÓt cependant dťpourvu d'activitť lipolytique. Peut Ítre la digestion des lipides a-t-elle lieu essentiellement chez ces espŤces dans le milieu intracellulaire aprŤs absorption des micelles lipidiques. Les hirudinťes paraissent ťgalement dťpourvues d'enzymes lipolytiques digestives d'origine endogŤne. La digestion des lipides impliquerait ici des enzymes bactťriennes.
Des sels biliaires sont prťsents chez tous les vertťbrťs depuis les agnathes. Chez ces derniers de mÍme que chez les sťlaciens, les tťlťostťens et les amphibiens, il s'agit essentiellement de sulfo-conjuguťs d'alcools biliaires tel le sulfate de scymnol ( figure 5-12) mis en ťvidence pour la premiŤre fois par Hammarsten dŤs 1898 (Hoppe-Seyl Z., 24, 322). Les conjuguťs d'alcools biliaires disparaissent ŗ partir des reptiles ŗ partir desquels on ne trouve plus que des conjuguťs d'acides biliaires. Les glyco-conjuguťs paraissent limitťs aux mammifŤres euthťriens.
On a ťgalement mis en ťvidences les propriťtťs ťmulsifiantes des sucs du tube digestif chez de nombreux invertťbrťs. Dans la plupart des cas, la nature des ťmulsifiants reste inconnue. Chez les crustacťs dťcapodes, un des composťs majeurs est un acide gras combinť avec de la taurine et de la sarcosine : la N-dodťcťnoyl-sarcosyl-taurine (figure 5-12).
Figure 5-12: Emulsifiants d'invertťbrťs et de vertťbrťs infťrieurs. A: Le sulfate de scymnol, un sulfo - conjuguť d'alcool biliaire prťsent chez les sťlaciens; le noyau de base cyclopentanophťnanthrťnique est le mÍme que celui des conjuguťs prťsents chez les mammifŤres (cfr. Figure 5-11). B: Le N – dodťcťnoyl – sarcosyl – taurine, un ťmulsifiant majeur des crustacťs dťcapodes.
Les ťtapes majeures de la digestion des protťines, des glucides et des lipides intervenant chez les mammifŤres sont rťsumťes dans le schťma de la figure 5-13. Cette figure reprend ťgalement les mťcanismes du contrŰle de la sťcrťtion des diffťrents sucs qui imprŤgnent le bol alimentaire. Dans ce cadre, la portion antťrieure, duodťnale, de l'intestin joue un rŰle particuliŤrement important. Outre les hormones sťcrťtine et pancrťozymine qui exercent des effets dťterminants essentiellement sur les sťcrťtions biliaires et pancrťatiques, le duodťnum produit ťgalement une sťrie de substances ŗ action hormonale ou locale tels l'entťrokinase, une enzyme activant le trypsinogŤne en trypsine, la motiline (ou villikinine), une substance favorisant la motricitť, le VIP (Vaso active Intestinal Peptide) qui a un effet vasodilatateur sur les capillaires irriguant l'intestin ou le GIP (Gastric Inhibitory Peptide) qui inhibe la production du suc gastrique. La portion terminale de l'intestin grÍle et le gros intestin produisent par ailleurs un PYY (peptide tyrosine - tyrosine) qui inhibe de nombreuses fonctions digestives (voir 4.2.2 ci-aprŤs).
Figure 5-13: La digestion des aliments chez les mammifŤres. G S: glandes salivaires pouvant produire des lipases et des amylases chez certaines espŤces, V B: vťsicule biliaire. Explications dans le texte (sections 2.2, 2.3 , 2.4 et aussi 4.1 et 4.2).
Les ťtapes de la digestion rťsumťes dans le schťma considŤrent uniquement des processus endogŤnes, propres ŗ l'organisme considťrť. Le tube digestif contient en gťnťral des micro-organismes en quantitťs plus ou moins importantes qui vont sťcrťter leurs propres enzymes digestives. Ces enzymes "exogŤnes" participeront de faÁon plus ou moins importante ŗ la digestion. Dans certains cas, les micro-organismes pourront par ailleurs servir d'apport de nourriture non nťgligeable. Nous allons maintenant envisager ces points particuliers plus en dťtail en considťrant les apports symbiotiques de nutriments.