 |
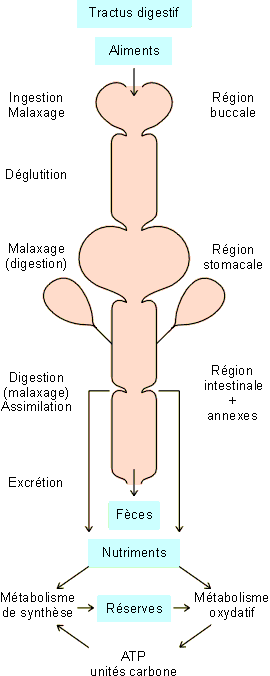
Figure 5-1: Le tube digestif et les fonctions de nutrition. |
1.2.1. Ingestion cellulaire ou extracellulaire
1.2.2. Microphages et macrophages, aliments liquides et solides
1.2.3. Quantité de nourriture
1.2.4. Qualité et nature de la nourriture
1.2.5. Phagostimulants et phagorepellants
1.5.1. La vessie gazeuse
1.5.2. Le jabot
En principe, la nutrition concerne l'ensemble des phénomènes d'échange impliqués dans la fourniture d'énergie métabolique et donc dans le maintien de l'activité cellulaire. Dans ce cadre, la respiration fait partie intégrante des fonctions de nutrition. Les physiologistes s'accordent cependant pour ne regrouper sous ce terme que les phénomènes intervenant dans les échanges de produits non gazeux, c'est-à-dire l'alimentation et l'excrétion. Par ailleurs, nombreux sont ceux qui, à l'heure actuelle, excluent l'excrétion des fonctions de nutrition, limitant donc celles-ci aux seuls processus d'alimentation.
L'alimentation concerne la prise de nourriture (ingestion), la digestion et l'assimilation. Dans ce processus, les aliments ingérés sont transformés par digestion en nutriments, éléments directement assimilables au niveau intestinal et utilisables par les différents organes dans le cadre de leurs activités (figure 5-1). L'essentiel des nutriments va être utilisé dans le métabolisme oxydatif qui fournira l'énergie nécessaire aux activités cellulaires (cfr. chapitre 3), au métabolisme de synthèse et à la constitution de réserves énergétiques.
Chez les animaux, le métabolisme de synthèse est incapable de fournir aux cellules l'ensemble des éléments qui leurs sont nécessaires. Ils doivent dès lors trouver par alimentation ces composés dits essentiels; les autres, dits non essentiels, seront synthétisés à partir des nutriments, soit au niveau de tous les tissus soit au niveau de certains organes seulement en utilisant l'énergie produite par le métabolisme oxydatif. Dans ce cadre, les animaux sont dits hétérotrophes par opposition aux organismes autotrophes (plantes, bactéries, par exemple) qui peuvent synthétiser l'ensemble des éléments dont ils ont besoin à partir de composés inorganiques en utilisant diverses sources énergétiques (solaire: plantes, bactéries hélioautotrophes - oxydation de composés minéraux : bactéries chimioautotrophes).
Les composés essentiels sont éminemment variables selon les espèces. Au sein d'une même espèce, ils peuvent parfois différer suivant le stade de développement ou l'évolution d'une maladie par exemple. On connaît ainsi le besoin en cholécalciférol (vitamine D3, voir chapitre 8 : hormones, calcium) du nourrisson humain alors que l'adulte peut en réaliser la synthèse en quantité suffisante.
Dans certains cas, des quantités appréciables de nutriments et de composés essentiels seront fournis par des bactéries, protozoaires ou algues symbiotiques (voir plus loin). Chez différentes espèces parasites, l'essentiel de la ration alimentaire consiste en nutriments fournis par l'hôte. Ces matières organiques dissoutes (MOD) paraissent pouvoir jouer un rôle significatif, quoique vraisemblablement toujours secondaire, chez différentes formes libres d'invertébrés aquatiques habitant des milieux où la charge organique est élevée. On a ainsi démontré la capacité de nombreux mollusques, crustacés et échinodermes à prélever des MOD (acides aminés, sucres) directement du milieu extérieur. Pour la plupart, les animaux auront cependant à prélever les aliments qui leurs conviennent dans la nature pour assurer leurs besoins.
1.2.1. Ingestion cellulaire ou extracellulaire
Dans la plupart des cas, l'ingestion implique la prise de nourriture au niveau d'une
cavité buccale. Les aliments y sont réduits en morceaux de petites tailles qui peuvent
être déglutis et conduits vers les parties du tube digestif assurant des fonctions
digestives. Chez différentes espèces primitives, l'ingestion s'effectue au niveau
cellulaire et implique une activité phagocytaire. Ainsi, chez bon nombre
d'unicellulaires, les particules alimentaires sont englobées au niveau de la membrane
plasmique dans des vésicules endocytaires et de là transférées dans les lysosomes où
s'effectue une digestion intracellulaire (voir plus loin et surtout
livre II, physiologie cellulaire). Chez les
cœlentérés et les plathelminthes, les particules entrent par une bouche dans un
tube digestif (cavité gastrique) sans différentiation dont l'épithélium est constitué
de cellules phagocytaires qui vont les ingérer. Chez les autres espèces le tube digestif
présente des différentiations plus ou moins importantes; son organisation est
éminemment variable suivant les espèces. Généralement, on peut néanmoins distinguer
trois parties essentielles (figure 5-1) : a) une partie buccale
impliquée dans la prise de nourriture et éventuellement dans son découpage en
"bouchées" qui puissent être avalées (bouchées déglutissables) ; b) une
partie stomacale reliée à la précédente par l'œsophage qui, dans la plupart des
cas, n'est qu'un simple conduit de liaison. Le développement de l'estomac et de
l'œsophage ainsi que leurs fonctions dans les processus nutritionnels sont
excessivement variables. Nous envisagerons certaines particularités de ces structures en
considérant les processus digestifs ; c) une partie intestinale souvent pourvue de
différentes annexes dont le rôle est essentiellement digestif.
1.2.2. Microphages et macrophages, aliments liquides et
solides
Bien que leurs aliments soient extrêmement variés, on peut regrouper les animaux en
différentes catégories en fonction de la nature et/ou de la dimension des particules
qu'ils ingèrent. On distinguera ainsi les consommateurs d'aliments liquides des
consommateurs d'aliments solides et, parmi ces derniers, les microphages des macrophages.
La consommation d'aliments liquides est le fait des animaux se nourrissant de nectar végétal (insectes, colibris,...), de sang (sangsues, tiques, moustiques, vampires,...) ou de lait (jeunes de mammifères). Une variante de ce type de prise d'aliment se retrouve chez les araignées et diverses larves d'insectes qui infusent des sucs digestifs dans les proies qu'elles capturent puis aspirent les tissus liquéfiés, plus ou moins digérés.
Les consommateurs d'aliments solides seront divisés en microphages et macrophages selon la dimension des particules alimentaires par rapport à leur propre taille.
Chez les animaux microphages, les particules alimentaires sont de petite taille et doivent être prises en masse dans le milieu. Les "particules" alimentaires vont ici de débris organiques de quelques nanomètres chez certains mollusques à des crevettes de relativement grande taille constituant une partie importante du "krill" dont se nourrissent certaines baleines et dont Euphausia superba est le type. Les systèmes de prise de nourriture sont variés : cils vibratiles assurant un courant d'eau et/ou mucus engluant les particules chez les mollusques et les tuniciers par exemple, structures de filtrage chez les crustacés, les flamants roses, les baleines, etc... Certaines espèces comme les lombrics ou les arénicoles ingèrent sans discernement la totalité du milieu qui les entoure et qui contient les particules alimentaires.
Chez les animaux macrophages, les aliments sont des éléments de grande taille prélevés individuellement dans le milieu. A l'inverse de ce qui se passe chez les microphages, les aliments sont ici choisis dans un processus mettant en jeu des systèmes sensoriels contrôlant la sélection et la prise de nourriture. La prise est généralement discontinue. Au niveau de la bouche, il y a souvent une étape de trituration et de mastication qui prépare la nourriture à être déglutie et transférée vers les autres parties du système digestif. Les cavités buccales présentent ainsi une grande variété de structures anatomiques permettant la prise correcte des aliments spécifiques et leur traitement avant déglutition. L'étude de ces appareils relève de la morphologie fonctionnelle plutôt que de la physiologie. Elle ne sera donc pas envisagée ici. Très brièvement, les structures impliquées dans la prise de nourriture sont nombreuses et très variées chez les invertébrés. Chez les vertébrés, elles sont moins nombreuses, se résumant au plus à deux mâchoires, une langue et des dents. Elles présentent cependant des particularités correspondant à des spécialisations dont l'étude, tout comme celle des pièces buccales des invertébrés sortent du cadre de ce traité.
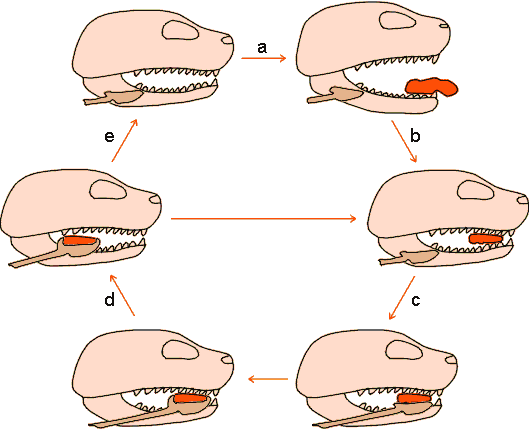
La prise de nourriture puis son transport vers l'œsophage implique un certain nombre d'étapes communes chez la plupart des vertébrés et en tout cas chez les tétrapodes (figure 5-2). La nourriture est saisie et serrée entre les mâchoires (a, b). Celles-ci s'ouvrent ensuite légèrement pour permettre à la langue de faire contact (c) puis éventuellement de prendre la nourriture en charge pour l'emmener vers l'arrière dans un mouvement de retrait concomitant à une phase d'ouverture rapide des mâchoires (d). Les mâchoires se referment ensuite, maintenant la nourriture, ce qui permet à la langue de se libérer et de revenir en place. La nourriture est ensuite avalée ou le cycle peut recommencer pour aboutir à une meilleure trituration et éventuellement à son transport jusqu'au pharynx où elle pourra finalement être déglutie et passer dans l'œsophage (e).
Figure 5-2: La prise de nourriture chez les vertébrés tétrapodes macrophages (explications dans le texte, d'après Kardong 1995, modifié).
Les aliments sont comme cela avalés entiers chez de nombreuses espèces. Ils peuvent aussi être triturés, déchirés, mâchés et mêlés à des sécrétions salivaires ayant ou non des propriétés digestives (voir 1.3 ci-après). Le rôle de la langue dans ce cas est de former une bouchée dite déglutissable et de l'amener à l'entrée du pharynx.
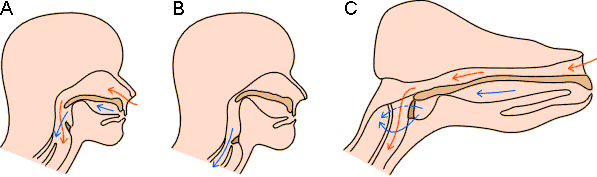
L'ingestion et la formation des bouchées déglutissables sont toujours des actes volontaires, la décision d'avaler ou de rejeter étant en rapport avec l'analyse de différentes informations sensorielles (voir 1.2.4 ci-après). Dans ce cadre les étapes c et d de la figure 5-2, au cours de laquelle la langue entre en contact avec la nourriture, jouent un rôle important. Une fois la nourriture "acceptée", le reste de la séquence s'effectue par voie réflexe. Sous la pression de la langue, le bol alimentaire est poussé dans le pharynx puis, par contraction des muscles du pharynx, dans l'œsophage. Chez bon nombre d'espèces, comme l'homme par exemple, ce processus peut couper momentanément la respiration: la glotte se ferme et l'os hyoïde ainsi que le larynx sont soulevés jusqu'à obturation complète des voies respiratoires (figure 5-3). Chez certaines espèces, l'organisation des voies aériennes supérieures et du début du tractus alimentaire est telle que la respiration peut être maintenue pendant toute la durée de la déglutition qui peut parfois être longue (serpents : voir chapitre 3; cétacés, opossum : voir figure 5-3). Une fois dans l'œsophage, le bol alimentaire est entraîné vers l'estomac par les ondes péristaltiques que provoquent les contractions des muscles entourant l'œsophage.
Figure 5-3 : Passage de l'air ( à) et de la nourriture ( à) chez les mammifères. Chez différentes espèces, l'homme notamment, l'organisation du pharynx place l'épiglotte relativement bas créant une possibilité d'ouverture de l'œsophage et de la trachée à la fois à l'air et à la nourriture (A). En fait lors de la déglutition, la respiration est momentanément suspendue. Sous la pression de la langue, poussant la nourriture vers l'arrière, l'os hyoïde ainsi que les parois du larynx sont déplacées jusqu'à obturation de la trachée par l'épiglotte et des voies respiratoires par l'os hyoïde (B, voir aussi figure 5-7). Chez d'autres espèces (C), le bol alimentaire peut passer autour de la trachée qui reste alors ouverte sur les conduits aériens.
Chez un même organisme, la nourriture ainsi ingérée peut varier largement en qualité et en quantité.
1.2.3. Quantité de nourriture
En principe, l'appétit, ou attrait pour la nourriture, intervient largement dans la
quantité de nourriture ingérée. Normalement, l'appétit égalise avec une bonne
précision les besoins énergétiques des organismes. Ceci paraît vrai chez la plupart
des espèces où ces problèmes ont pu être abordés, vertébrés comme invertébrés.
L'essentiel des études effectuées jusqu'à présent porte sur les vertébrés
supérieurs. Chez eux, la prise de nourriture paraît essentiellement contrôlée par
l'activité des centres hypothalamiques de la faim et de la satiété. Ces centres
intègrent des stimuli centraux et périphériques positifs ou négatifs qui vont stimuler
ou inhiber l'attrait pour la nourriture. Certains de ces stimuli sont à action rapide,
déterminant une variation d'appétit "immédiate". D'autres sont à action
nettement plus lente, régulant la prise d'aliments sur des périodes qui se comptent en
semaines, en mois voire en années.
Le taux de glucose sanguin est l'un des stimuli à action rapide les plus importants. Une diminution de la glycémie sanguine déterminera très rapidement une sensation de faim alors qu'une augmentation induira une inappétence marquée.
La distension gastrique apparaît également comme un stimulus négatif à action rapide important. C'est lui qui déterminera essentiellement la sensation de satiété qui, à la fin d'un repas, arrêtera la prise de nourriture. De même certaines hormones de la digestion comme l'entérokinase (voir plus loin) paraissent avoir un effet rapide sur le centre de la satiété, du moins chez le rat. Il en va de même pour la cholecystokinine (= pancréozymine, voir plus loin) dont l'effet comme agent de satiété a été mis en évidence chez de nombreux vertébrés mais qui pourrait être active aussi chez les invertébrés. Ainsi, chez le mollusque Nivanax inermis, un composé de type cholecystokinine paraît inhibé l'activité des neurones des ganglions buccaux contrôlant la capture des proies (Zimering, Madsen et Elde, Peptides 9, 133, 1988).
Diverses substances de nature opioïdes (les kappa opioïdes) ont par ailleurs été décrites comme stimulant la prise de nourriture chez les vertébrés mais aussi chez quelques invertébrés. L'essentiel des études actuelles porte sur les effets du naloxone , un antagoniste de ces opioïdes (Mosley, Am. Zool. 35, 437, 1995). Le naloxone agit comme un puissant dépresseur d'appétit chez bon nombre d'invertébrés et de vertébrés. Le mode d'action des opioïdes reste à l'heure actuelle inconnu.
Le niveau des réserves lipidiques et le taux d'acides gras libres dans le sang apparaissent comme des facteurs importants de régulation à long terme. Certains résultats récents indiquent un contrôle via une "hormone", la protéine Ob, du niveau des réserves lipidiques ainsi que de l'importance des entrées alimentaires (cfr. 6.4 ci-après).
L'état psychique (stress, maladie), de même que la température corporelle peuvent également intervenir et devenir, le cas échéant, des facteurs prépondérants, à court comme à long terme.
Chez l'homme et d'autres espèces sociales, d'autres paramètres peuvent intervenir de façon importante dans les comportements nutritionnels (habitudes, mœurs, mode ...).
Grâce à ces mécanismes et à d'autres, encore très mal connus actuellement, les organismes peuvent ajuster leur prise de nourriture à leurs besoins et conserver ainsi une quantité relativement constante de tissus adipeux (graisses de réserve, voir plus loin). Une prise de poids anormale (obésité) paraît due à une défaillance d'un ou plusieurs systèmes impliqués dans ce processus homéostatique. Peu de choses sont actuellement claires à ce sujet. Certaines espèces paraissent plus sujettes à dérèglement que d'autres.
1.2.4. Qualité et nature de la nourriture
Il n'existe que peu de données physiologiques précises sur la sélection de la
nourriture chez les animaux. Comme nous l'avons déjà signalé, la prise de nourriture
est toujours, chez toutes les espèces, un acte volontaire et toute une série de
paramètres vont intervenir dans la décision d'avaler ou de rejeter une nourriture. Ces
paramètres sont nombreux; ils relèvent de facteurs anatomiques génétiquement fixés,
de stimuli nerveux en rapport avec les qualités olfactives, gustatives ou visuelles de la
nourriture, de facteurs comportementaux innés ou acquis.
Les facteurs morphologiques vont clairement limiter la sélection possible des aliments. Comme nous l'avons déjà signalé, les systèmes de prise de nourriture sont chez les animaux nombreux et variés et certains types de structures excluent d'office certains types d'aliments (un léopard n'est pas bâti pour se nourrir comme un colibri du nectar des fleurs ! - voir aussi 1.2.3 ci-avant).
Les autres facteurs vont intervenir de façon très complexe dans le cadre d'une intégration nerveuse dont sortira la décision de rejeter ou d'avaler. Les mécanismes gérant cette intégration sont actuellement très mal connus et font intervenir de très nombreux facteurs. Chez bon nombre d'espèces, la présence dans les aliments de phéromones, de poisons ou de toxines constituent des paramètres d'importance dans la prise de décision.
1.2.5. Phagostimulants et phagorepellants
Bien qu'il ne vienne à l'esprit de personne de nier l'existence de composés dans les
aliments ayant des effets directs sur l'ingestion chez les vertébrés y compris chez
l'homme, ces substances ont été essentiellement étudiées chez les invertébrés. Un
des premiers exemples fut celui des interactions feuille de mûrier - vers à soie
étudié dès 1962 par une équipe japonaise (Hamamura et al, Nature 194,
754, 1962).
Il ressort de ces expériences qu'une série de composés présents dans les feuilles du mûrier induisent chez la chenille du Bombyx mori tout un comportement en relation avec la prise de nourriture (tableau 5-1). Ainsi, la chenille est attirée vers les feuilles de mûrier et commence à les mordre par un mélange de monoterpènes : acétate de terpinyl et de linalyl, linalol, citral et b - g - hexenol. La mastication sera entretenue par la présence de sucres (inositol, saccharose) de terpenoïdes (sitosterol) et de flavonoïdes (isoquercitrine, morine). La déglutition sera stimulée notamment par la présence de cellulose. Certaines de ces substances sont extrêmement spécifiques; leur absence ou simplement une légère modification de structure peut bloquer le comportement nutritionnel. Ainsi, la substitution de l'unité glucose de l'isoquercitrine par du rhamnose (figure 5-4) inhibe totalement la mastication.
Tableau 5-1
Facteurs des feuilles de mûriers associés à la prise de nourriture chez le vers à
soie
| Attractants | Facteurs de mastication | Facteurs de déglutition |
| Citral | Sucrose | Cellulose |
| Acétate de terpinyl | Inositol | Silicate |
| Acétate de linalyl | Sitosterol | Phosphate |
| Linalol | Isoquercitrine | |
| b - g - hexenol | Morine |
Depuis ces premières études, les recherches se sont multipliées et les exemples de composés pouvant stimuler ou inhiber la prise de nourriture chez les invertébrés se sont multipliés. Eminemment variables suivant les espèces, ils peuvent agir via les systèmes gustatifs ou olfactifs. Une étude détaillée de ces composés sort du cadre de ce traité.
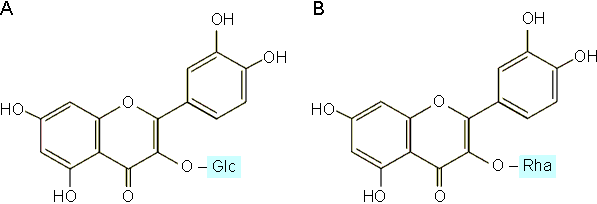
Figure 5-4: Isoquercitrine stimulant la mastication (A) chez le ver à soie et son dérivé la bloquant complètement (B).
A côté de ces produits, différentes substances dites de défense (toxines, venins) présentent dans des organismes pouvant servir de nourriture peuvent avoir un effet inhibiteur marqué direct puis indirect, après apprentissage. Souvent d'ailleurs, les organismes possédant ce genre de toxines se rendent facilement repérables par signaux visuels ou olfactifs pour éviter de servir de proie.
Les substances appartenant à ce groupe sont également nombreuses et variées tant chez les végétaux que les animaux. Elles vont des quinones aux peptides neurotoxiques en passant par des acides, des terpènes, des phénols ou des alcaloïdes (voir par exemple Harborne, 1977 ou Goyffon et Heurtault, 1995). Elles vont essentiellement intervenir dans une sélectivité par apprentissage de la nourriture. Une catégorie bien connue de toxiques végétaux est celle des alcaloïdes tels que l'atropine, la solanine ou la strychnine dont les effets sur le système nerveux des vertébrés sont bien connus (voir livre 2, physiologie cellulaire). Une autre catégorie bien connue est celle des glycosides cyanogéniques. Ces composés, que l'on retrouve par exemple dans différentes graines (odeur caractéristique: amande amère), libèrent de l'acide cyanhydrique par hydrolyse suivant le schéma général :

Les animaux faisant l'expérience de ce type de plantes en doses non létales cessent en général d'en ingérer et apprennent par ailleurs très vite à les reconnaître.
Dans le cas des plantes à glycosides cyanogéniques, on peut cependant noter l'acquisition d'une certaine résistance due à la synthèse d'une enzyme de détoxification, la rhodanèse, transformant le cyanate en sulfocyanate à partir d'acide b - mercaptopyruvique. Cette résistance a été mise en évidence sur des troupeaux de moutons mais aussi chez l'homme, dans le cas de tribus africaines se nourrissant de farine de racines de cassava.
Certains organismes apprennent par ailleurs à se servir de toxines produites par d'autres pour éloigner les prédateurs. Ainsi par exemple, les plantes à latex de la famille des Asclépiadacées produisent des glycosides cardiaques essentiellement toxiques pour les vertébrés supérieurs. L'asclépiade de Curaçao (A. curassavica) est ainsi devenue une des nourritures favorites de la chenille d'un papillon monarque Nord américain qui se gorge de glycosides, notamment la calotropine, les transmets au papillon que les oiseaux apprennent très vite à reconnaître.
Chez bon nombre d'espèces macrophages, la formation de bouchées déglutissables implique une lubrification et parfois une humidification (espèces terrestres) de la nourriture ingérée. Les glandes salivaires sont généralement absentes chez les poissons et les batraciens. On trouve dans la cavité buccale de ces espèces des cellules ou des groupes de cellules sécrétant un mucus aidant à la déglutition. Les glandes salivaires sont également absentes chez la plupart des reptiles. Chez certaines espèces cependant les cellules à mucus sont organisées en glandes plus ou moins complexes. Les oiseaux possèdent en général des glandes salivaires bien développées. Elles sécrètent cependant essentiellement du mucus et peu de liquide. Il n'y a en définitive que chez les mammifères que les glandes salivaires produisent des quantités de liquide appréciables. Cette caractéristique est vraisemblablement liée au fait que les mammifères, à l'inverse des autres vertébrés, réduisent leur nourriture en bouche à un état plus ou moins pâteux. Cette faculté permet une déglutition plus aisée mais aussi une analyse plus fine des particularités chimiques des aliments ingérés. Chez les autres vertébrés, la nourriture est le plus souvent avalée telle qu'elle ou après une trituration et une mise en pièce sommaire.
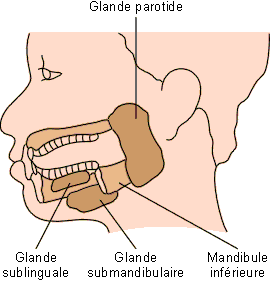
Chez les mammifères, la salive résulte essentiellement de l'activité de trois formations glandulaires : les glandes parotides se trouvant à l'arrière de la bouche, les glandes sublinguales et submandibulaires (sous-maxillaires ou submaxillaires) se trouvant sous le plancher buccal (figure 5-5).
Il existe des variations considérables de volume et de localisation précise des glandes salivaires chez les différents groupes de mammifères. Chez les cétacés et autres mammifères marins, leur volume est en général extrêmement réduit. Chez de nombreux édentés, comme le tamanoir par exemple, ce sont les glandes submaxillaires qui sont de loin les plus développées, elles sont même munies d'une vessie de stockage. Chez les ongulés par contre, les glandes parotides sont souvent bien développées. Elles produisent une salive essentiellement aqueuse, servant à diluer l'énorme bol alimentaire de ces herbivores (voir plus loin : digestion de la cellulose). Les autres glandes produisent des liquides généralement plus riches en mucines.
La production de salive est continue. Chez l'homme, elle s'élève en moyenne à 1500 ml/24h ou 1 ml/min environ dont 70 % proviennent des glandes submandibulaires (sous-maxillaires), 25 % des parotides et les 5 % restants des glandes sublinguales ainsi que de formations mineures de la muqueuse buccale. Chez les ruminants, la production salivaire peut aller de 5 à 15 litres par jour chez les moutons et les chèvres et jusqu'à 200 litres par jour pour le bétail de grande taille. La production de salive est éminemment variable en fonction de la disponibilité de l'eau et de la balance hydrique. En cas de carence en eau, la bouche et la gorge deviennent rapidement sèches, ce qui peut rendre la déglutition difficile.
Les rôles majeurs des sécrétions salivaires sont la lubrification et l'humidification. Les glandes peuvent également remplir d'autres fonctions. On notera ainsi, suivant les espèces : sécrétion de venins et de toxines, sécrétion d'enzymes digestives (amylase, voir plus loin) - sécrétion d'anticorps et d'enzymes bactériostatiques assurant une certaine protection contre les micro-organismes - sécrétion de HCO3- alcalinisant et procurant un certain pouvoir tampon à la salive. On a également mis en évidence chez les mammifères, y compris l'homme, la sécrétion d'une hormone de croissance qui accélère la guérison des blessures léchées.
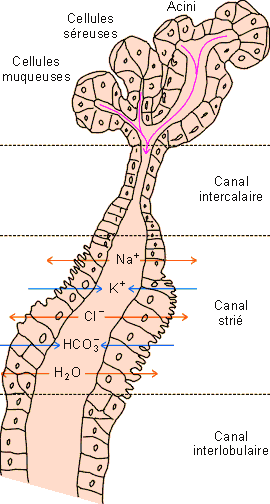
Les acini des glandes salivaires (figure 5-6) représentent le lieu de production de la salive; la salive primaire, qui y est formée, a une composition en électrolytes et une osmolarité voisine de celle du plasma sanguin. Elle subit des modifications lors de son passage dans les canaux excréteurs. Du Na+ et du Cl- sont réabsorbés alors que du K+ du HCO3- et d'autres ions (I-, F- notamment) y sont sécrétés (mécanismes de transport : cfr. livre 2, physiologie cellulaire). La sécrétion est en général largement hypo - osmotique par rapport au plasma sanguin (±70 mOsm/kg H2O chez l'homme au repos contre ±300 mOsm/kg H2O pour le plasma). Son osmolarité varie largement avec la quantité produite. Lorsque la salivation augmente, le passage au niveau des conduits modificateurs s'accélère, diminuant ainsi la réabsorption de Na+ et de Cl- ainsi que la concentration de K+ et de HCO3- ( figure 5-6). Différents types cellulaires sécrètent par ailleurs les mucines et les protéines que contient la salive. La production de salive est réflexe, sous contrôle d'un centre salivaire bulbaire. Elle dépend des systèmes sympathique et parasympathique. Les activations cholinergique, a - adrénergique et peptidergique stimulent la formation d'une salive aqueuse alors que l'activation b - adrénergique conduit à une salive très visqueuse riche en mucines et protéines. La salivation peut par ailleurs être largement augmentée par différents stimuli tels que l'odeur et le goût des aliments ou la mastication.
Comme nous l'avons déjà signalé, la déglutition est un acte volontaire au cours duquel les bouchées dites déglutissables sont poussées par mouvements musculaires, notamment de la langue chez les tétrapodes, vers l'entrée de l'œsophage. Le bol alimentaire est ensuite poussé vers l'estomac par des mouvements péristaltiques ( figure 5-7). Rappelons également que chez de nombreuses espèces, le passage de la nourriture dans le pharynx bloque momentanément la respiration.
Figure 5-7: La déglutition chez l'homme : la bouchée déglutissable du bol alimentaire est poussée vers l'œsophage puis vers l'estomac par des contractions des muscles de la langue, du pharynx puis de l'œsophage.
L'œsophage est essentiellement un conduit amenant la nourriture de la bouche à l'estomac; on n'y trouve aucune structure particulière. Il est tapissé d'un épithélium stratifié squameux au niveau duquel s'ouvrent des glandes sécrétant un mucus qui facilite le passage du bol alimentaire. Il est entouré de couches musculaires lisses dont le péristaltisme pousse la nourriture vers l'estomac. L'entrée au niveau stomacale est normalement fermée par un sphincter (cardia) dont l'ouverture est réflexe et peut être commandée par le passage du bol alimentaire au niveau du pharynx. La déglutition entraîne également, par stimulation de fibres vagales, un relâchement des fibres musculaires de l'estomac proximal ce qui permet une certaine distension et la réception du bol alimentaire venant de l'œsophage (réflexe de relaxation réceptive). C'est au niveau de l'estomac que débute chez bon nombre d'espèces le processus de digestion qui est brièvement envisagée dans la section 2 ci-après.
Dans certaines conditions, le mouvement du bol alimentaire peut être inversé, allant de l'estomac vers la bouche; le vomissement est en général un phénomène réflexe pouvant faire intervenir de nombreux facteurs : remplissage trop important de l'estomac, inflammation de la muqueuse gastrique, irritation des organes d'équilibration (cinépatie), pression sur le cerveau, stress important, douleurs violentes, odeurs ou visions écœurantes et contacts avec la muqueuse pharyngienne constituent autant de facteurs d'activation du centre de vomissement situé dans le bulbe rachidien, au niveau de la formation réticulée.
Chez les tétrapodes, le vomissement implique des contractions brusques et importantes des muscles abdominaux; ce qui a pour effet de comprimer l'estomac et de pousser le contenu gastrique vers l'œsophage, le sphincter cardiaque étant ouvert. Les rats et les chevaux sont incapables de vomir. Les rats sont en général extrêmement sélectifs en ce qui concerne la qualité de leur nourriture de façon à éviter tout problème. Les chevaux le sont malheureusement moins et le réflexe de vomissement provoque souvent chez eux une déchirure de l'estomac. On met en général en cause dans ce problème un sphincter et une plicature musculaire cardiaque très puissants qui empêchent la remontée du bol alimentaire (voir plus loin).
Chez de nombreuses espèces (oiseaux, mammifères notamment), la régurgitation du bol alimentaire peut être contrôlée dans le cadre de la nutrition des jeunes ou de la rumination (voir plus loin). La régurgitation est accompagnée d'un antipéristaltisme de l'œsophage ce qui ne paraît pas être le cas du vomissement.
L'œsophage donne naissance chez certains animaux à deux types de différentiations particulières.
1.5.1. La vessie gazeuse
Chez beaucoup d'actinoptérygiens, la paroi dorsale de l'œsophage comporte un
diverticule impair se développant vers l'arrière en un sac volumineux, immédiatement
sous la colonne vertébrale, entre les reins. Cette structure, appelée vessie gazeuse
reste, dans la plupart des cas, en communication avec l'œsophage par un canal dit
pneumatique. Cette connexion disparaît chez les téléostéens les plus évolués. La
vessie gazeuse contient un gaz de composition variable suivant les espèces mais jamais
semblable à celle de l'air atmosphérique. Il est en général assez riche en oxygène
(88 % chez la carpe par exemple). La charge en oxygène dépend de mécanismes complexes
d'échange pouvant mettre en jeu des propriétés particulières des hémoglobines (voir
chapitre 4). Son rôle majeur reste peu clair à l'heure
actuelle. Initialement, on l'a considéré comme une aide à la vie aquatique, diminuant
la densité des poissons qui la possèdent, d'où son ancien nom de vessie natatoire. Ce
rôle apparaît maintenant comme assez secondaire. Chez certaines espèces d'eaux douces
peu oxygénées, la vessie gazeuse peut intervenir dans les échanges respiratoires. Nous
avons envisagé ce problème au chapitre 3 (3.3.1.c). La
vessie gazeuse est par ailleurs souvent en relation avec l'oreille interne. Certains
supposent que cette disposition augmenterait la sensibilité auditive des poissons.
1.5.2. Le jabot
Chez certains oiseaux, l'œsophage se dilate en avant de la ceinture pectorale en un
jabot; poche ventrale qui sert de réservoir où peuvent être stockés les aliments. Chez
les colombiformes, le jabot contient deux plages épithéliales glandulaires dont la
prolifération est activée pendant les derniers jours de l'incubation des œufs. La
desquamation de cet épithélium après l'éclosion, fournit une masse laiteuse qui
intervient dans la nourriture des poussins jusqu'au 15ème jour (lait de
pigeon). La prolifération de l'épithélium est stimulée par la prolactine, une hormone
stimulant la sécrétion lactée chez les mammifères (voir chapitre
9 sur les relations hormonales).