3.3.1. Les systèmes de respiration aérienne
a. cavités buccales
b. cavités pharyngiennes et branchiales
c. vessie gazeuse
d. système gastro-intestinal
e. poumons3.3.2. Les échanges gazeux
3.4.1. Les poumons des vertébrés terrestres
a. les poumons alvéolaires
b. les poumons tubulaires3.4.2. Le système trachéen des insectes
3.4.3. Conduits ventilatoires, échanges hydriques et thermiques
3.4.4. La ventilation et son contrôle
a. mouvements et mécaniques ventilatoires
b. cycles et fractions respiratoires
c. régulation de la respiration1. Les centres respiratoires
2. Les stimuli mécaniques et la rythmicité
3. Les stimuli chimiques et l'adaptation de la ventilation
4. Autres stimuli
Le milieu aérien est nettement plus riche en oxygène que le milieu aqueux et bon nombre d'animaux essentiellement aquatiques dont la teneur en oxygène peut devenir basse ont appris à utiliser l'air atmosphérique pour obtenir le complément d'oxygène dont ils ont besoin. Cette respiration aérienne d'appoint se retrouve aussi bien chez des invertébrés que chez des vertébrés aquatiques.
3.3.1. Les systèmes de respiration aérienne
Dans le système le plus simple, une bulle d'air "happée" par
l'animal va augmenter la PO2 au niveau du liquide passant sur les
structures branchiales. Ce système est pratiqué par différents mollusques
intertidaux par exemple. Lorsque ces espèces sont exondées, elles s'isolent du
milieu extérieur par différents moyens (fermeture de valves, d'opercule, se
collent au substrat), empêchant ainsi une perte de liquide paléal mais aussi
tout échange avec l'extérieur. Le liquide paléal isolé s'appauvrit très
vite en oxygène. Certaines espèces peuvent enrichir ce milieu en O2
en prenant rapidement de petites quantités d'air à l'extérieur. Ce système simple est
aussi utilisé par certains poissons (carpes par exemple) lorsque la tension en
oxygène du milieu diminue. De même, le concombre de mer Holothuria tubulosa,
vivant dans des eaux stagnantes et peu profondes pauvres en oxygène, peut
happer de l'air au niveau de son cloaque où se trouve les branchies en
étendant sa partie arrière vers la surface.
Dans un système plus élaboré, les parois d'une ou de plusieurs cavités deviennent très vascularisées favorisant les échanges gazeux au niveau de l'air happé. Dans certains cas, il s'agira des cavités branchiales, buccales ou pharyngées, dans d'autres cas de la cavité intestinale ou encore de la vessie gazeuse. La peau peut aussi devenir un organe de respiration aérienne d'appoint notable chez certains poissons. Ainsi l'anguille commune qui peut parcourir de longues distances par temps humide pour aller d'une rivière à une autre. Il semble que près de 60 % de la prise d'oxygène s'effectue dans ces conditions par la peau.
Chez de nombreuses espèces la pratique de la respiration aérienne d'appoint n'a rien d'accidentel. Elle fait partie intégrante du mode de vie des animaux qui vont respirer tantôt dans l'eau, tantôt dans l'air. On parlera dans ces cas de respiration bimodale. La respiration bimodale fait toujours intervenir des organes particuliers impliqués spécifiquement dans la respiration aérienne. Ces organes sont en relation soit avec la cavité buccopharyngienne, la cavité branchiale ou le tube digestif. Elle est spécialement pratiquée par des espèces vivant dans des milieux dont la teneur en O2 peut être très basse.
a. cavités buccales
Le périophtalme des mangroves, commun dans les mangroves et dans les estuaires
boueux des régions tropicales, passe l'essentiel de son temps à l'air,
supporté par ses nageoires pectorales. La respiration aérienne s'effectue ici
dans la cavité buccale, hérissée de nombreuses et volumineuses papilles
richement vascularisées. L'anguille électrique (gymnote) des rivières
d'Amérique du sud utilise également sa cavité buccale dans un processus
respiratoire. Elle vient obligatoirement et régulièrement en surface (toutes les
unes à deux minutes) pour renouveler sa provision d'air.
b. cavités pharyngiennes et branchiales
Chez d'autres espèces de poissons, la respiration aérienne s'effectue dans des
diverticules pharyngiens ou operculaires. L'amphipnous, poisson apode et
amphibie des marécages de l'Inde possède deux évaginations dorsales de la
cavité pharyngienne qui lui servent de "poumons". De nombreuses
espèces de poissons-chats d'Afrique, d'Asie ou d'Amérique ont des chambres
operculaires très vascularisées. Le Monopterus asiatique possède ainsi
deux cavités suprabranchiales capables de se dilater en deux sortes de petits
ballons apparaissant en arrière de la tête. On retrouve ce genre de
diverticules chez l'Anabas, perche grimpeuse indienne. Chez l'Hétérobranchus
africain, la cavité suprabranchiale renferme des organes arborescents dérivés
de branchies, très ramifiés et vascularisés servant aux échanges gazeux (figure 3-33). Chez Saccobranchus, poisson-chat du
sud-est asiatique, les organes suprabranchiaux de respiration aérienne
s'allongent largement en arrière de la tête en deux sacs alvéolés. Il s'agit
là de véritables "poumons" operculaires inclus à l'intérieur des
myotomes du tronc et de la moitié antérieure de la queue.
Chez différents crustacés décapodes, amphibies ou terrestres, les parois de la cavité branchiale peuvent également contenir des organes de respiration aérienne. L'épithélium de la lame branchiostége s'organise ici en expansions tissulaires plus ou moins ramifiées, formant des "poumons" branchiostégiaux. Chez les crabes de la famille des ocypodidæ, les poumons sont peu développés; la respiration aérienne s'effectue essentiellement à partir de "fenêtres aériennes" (gas windows) se trouvant sur des segments aplatis des pattes. La cuticule au niveau de ces fenêtres est très mince (0,5 µm) et recouvre un système de capillaires relativement dense dont l'organisation ressemble à celle existant dans les poumons branchiostégiaux (figures 3-34A et 3-34B).
c. vessie gazeuse
Chez certaines espèces de poissons, la vessie gazeuse peut être impliquée
dans les échanges respiratoires. Chez la plupart des actinoptérygiens, la
paroi dorsale de l'œsophage émet un diverticule impair qui se développe vers
l'arrière en un sac ovale, s'insinuant dans le mésentère, entre les reins,
immédiatement sous la colonne vertébrale. Chez quelques holostéens et
téléostéens d'eaux douces vaseuses et peu oxygénées, l'épithélium de la
vessie gazeuse se soulève en replis richement vascularisés conférant à la
muqueuse une structure alvéolaire, de type pulmonaire. Chez Erythrinus,
un téléostéen d'Amazonie, ce poumon impair peut couvrir jusqu'à 60 % des
besoins en O2. Chez cette espèce, l'usage de ce système reste
facultatif. Il devient obligatoire chez le pirarueu (Arapaina gigas),
autre poisson amazonien qui est incapable de vivre sous eau et doit faire
surface très fréquemment comme le gymnote pour prendre de l'oxygène.
d. système gastro-intestinal
Chez d'autres espèces encore, la respiration aérienne sera assurée par
différentes parties du système gastro-intestinal. Différents poissons-chats
des marécages d'Amérique du Sud peuvent ainsi accumuler de l'air dans leur
estomac ou leur intestin, en partie transformé en une poche à air richement
vascularisée et dépourvue de toute fonction digestive. Ce genre de poche
respiratoire intestinale se retrouve chez la loche de nos étangs. L'air est
introduit soit par la bouche (poissons-chats) soit par l'anus (loches).
e. poumons
Les poumons ne sont pas l'attribut exclusif des seuls vertébrés terrestres. On
en trouve effectivement chez certains poissons actuels, coexistant avec
l'appareil branchial. On peut donc penser que le poumon est apparu au cours de
l'évolution bien avant la conquête des continents.
On trouve un appareil pulmonaire chez une quinzaine d'espèces de poissons appartenant à trois groupes anciens d'osteichthyens : brachioptérygiens, dipneustes et crossoptérygiens. Brachioptérygiens et dipneustes sont des poissons d'eaux douces de régions tropicales vivant dans des marécages ou des rivières soumis à des assèchements ou à fort appauvrissement en oxygène en saison sèche. Ces animaux estivent le plus souvent dans la boue et la respiration pulmonaire supplée alors la respiration branchiale devenue impraticable. Les crossoptérygiens sont des formes marines à poumons dégénérés inutiles mais représentant probablement la relique de poumons fonctionnels chez des formes primitives du Dévonien, ancêtres des tétrapodes.
Les poumons sont en général ici de simples sacs à parois lisses richement vascularisées. Ils sont dits unicavitaires ou sacculaires. Chez certains dipneustes, la cavité pulmonaire est toutefois divisée superficiellement par des septa de 1er ou de 1er et de 2ème ordre (voir plus loin: batraciens). L'épithélium respiratoire est fait d'un seul type cellulaire ayant les caractéristiques générales des pneumocytes de type 1 des mammifères mais contenant les corps multilamellaires osmiophiles des pneumocytes de type 2 (voir plus loin).
En fonction de l'espèce on trouvera un ou deux poumons dorsaux ou ventraux. Ces poumons s'ouvrent toujours au niveau de l'œsophage par une glotte munie d'un sphincter.
L'inspiration est assurée, comme chez les amphibiens, par compression du plancher buccal qui force l'ouverture de la glotte. L'expiration résulte de la contraction de muscles lisses entourant les poumons.
La respiration aérienne n'assure chez les poissons pulmonés que quelque 10 % des besoins en oxygènes normaux. Elle permet néanmoins la survie pendant la saison sèche lorsque les animaux estivent et qu'ils ont un métabolisme très bas.
On retrouve des poumons unicavitaires chez les amphibiens, un groupe pratiquant largement la respiration bimodale (poumon - peau) et restant largement inféodé aux milieux aquatiques ou très humides. La plupart des urodèles ont des poumons lisses. Les anoures et les apodes ont des poumons septés (figure 3-35). Comme chez les poissons, l'épithélium ne comporte qu'un seul type de pneumocytes cumulant les caractéristiques des types 1 et 2 des amniotes.
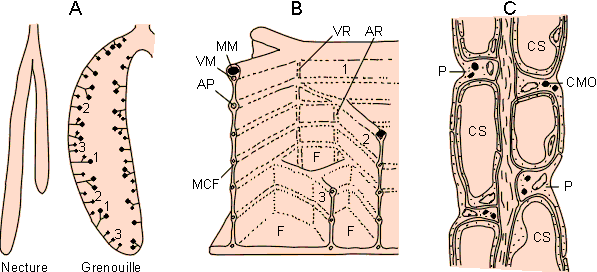
Figure 3-35: A: Poumons lisses et septés des batraciens. B: Organisation des septa montrant le cloisonnement en favéoles (F). C: Organisation d'un septum, essentiellement constitué de pneumocytes (P) contenant des corps multilamellaires osmiophiles (CMO) et entourant l'endothélium des capillaires sanguins (CS). AP: artère pulmonaire, AR: artère radiale, MCF: muscle circumfavéolaire, MM: muscle marginal, VM: veine marginale, VR: veine radiale. D'après Beaumont et Cassier 1987, modifié.
3.3.2. Les échanges gazeux
Il y a lieu de distinguer ici les échanges d'O2 de ceux de CO2.
Comme nous l'avons déjà signalé, l'élimination de CO2 est
beaucoup plus facile en milieu aquatique qu'en milieu aérien (HCO-3).
La prise d'oxygène sera par contre plus aisée en milieu aérien qu'en milieu
aquatique (concentration en O2). Par ailleurs, les structures
respiratoires d'appoint restent peu évoluées et n'ont guère l'efficacité des
poumons d'amniotes dans l'élimination du CO2. Dans de nombreux cas,
la respiration aérienne va dès lors pouvoir contribuer de manière
significative à l'apport en O2, elle ne va par contre intervenir que
de façon peu satisfaisante dans l'élimination du CO2. La figure 3-36 illustre ce phénomène dans le cas du
poisson-chat Symbranchus marmoratus. On remarque effectivement une
augmentation rapide de la teneur en O2 du sang dès que l'animal à
accès à l'oxygène de l'air. La teneur en CO2 sanguine augmente
cependant elle aussi, traduisant un défaut d'élimination lorsque l'animal est
dans l'air. Le CO2 diminue par contre dès que l'animal reprend une
respiration branchiale. Une bonne part du CO2 va s'éliminer dans ce
cas sous forme de HCO-3 (voir
chapitre 1, 5.4.4). L'accumulation de CO2
provoque une acidose sanguine (cfr. chapitre 4)
qui peut prendre pas mal de temps pour être contrôlée. Ainsi l'anguille
mettra-t-elle environ deux heures pour régulariser son pH sanguin après une
incursion de une heure en milieu aérien. La problématique est la même chez
les poissons pulmonés et bon nombre de batraciens où l'élimination de CO2
se fait toujours assez largement par voie aquatique alors que la voie aérienne
peut contribuer très largement à l'apport en O2 (figure
3-37).
Figure 3-36: Evolution de l'O2
et du CO2 artériel chez Symbranchus marmoratus respirant
dans l'air ou dans l'eau. Au temps 0, marqué par la flèche, l'animal gonfle
son pharynx, inspirant de l'air frais. D'après Johansen (Comp. Biochem. Physiol.,
18, 388, 1966), modifié.
 |
Figure 3-37: Contribution des voies aériennes et aquatiques aux échanges O2 - CO2 chez trois espèces de poissons pulmonés. D'après Lenfant, Johansen et Hanson (Fed. Proc. 29, 1124, 1970), modifié. |
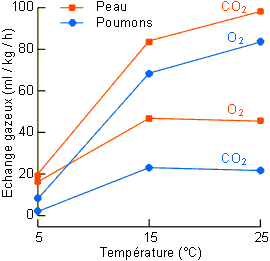
L'importance relative des voies aériennes et aquatiques dans les échanges gazeux va évidemment varier en fonction de l'animal considéré et, chez un même animal, en fonction de son activité métabolique. Dans certains cas, la respiration aérienne peut s'avérer suffisante pour couvrir les besoins d'O2 d'un organisme dans certaines conditions et pas dans d'autres. Ces conditions sont en général liées à l'activité métabolique et à la demande énergétique: repos - activité, variation de température chez les pœcilothermes (figure 3-38).
|
|
Le passage à la respiration strictement aérienne va dépendre 1) de la mise au point de surfaces d'échanges suffisamment importantes et efficaces pour pouvoir régler le problème de l'apport en O2 mais surtout comme nous l'avons vu plus haut, celui de l'élimination du CO2 2) d'adaptations en rapport avec la déshydratation de surfaces d'échanges importantes et largement ventilées 3) d'adaptations en rapport avec une augmentation significative de la teneur en CO2 sanguine.
Au plan respiratoire, on retrouve des systèmes d'échanges par voie aérienne efficaces aussi bien chez les invertébrés que chez les vertébrés. Il s'agit toujours de structures cavitaires de type "poumon" sauf chez les insectes qui utilisent un système trachéen. Chez les invertébrés, les espèces pulmonées sont de petite taille avec un métabolisme peu élevé (mollusques gastéropodes, scorpions ...). Les poumons ne sont guère performants; n'étant souvent pas spécifiquement ventilés ni particulièrement irrigués. Il n'existe à l'heure actuelle que peu de données physiologiques concernant ces structures. Nous n'en discuterons donc pas plus longuement.
3.4.1. Les poumons des vertébrés terrestres
Les poumons des premiers vertébrés réellement terrestres ne se distinguent
guère des poumons des amphibiens. Chez la plupart des reptiles en effet les
poumons sont unicavitaires avec des parois présentant des septa de ler
et 2ème ordre. On voit apparaître pour la première fois les deux
types de pneumocytes 1 et 2, caractéristiques des amniotes. Les pneumocytes de
type 1 sont essentiellement respiratoires alors que la sécrétion de
surfactants et de mucus est dévolue aux pneumocytes de type 2 (voir plus loin).
a. les poumons alvéolaires
Chez les chéloniens, les crocodiliens et les varanidés, les poumons deviennent
pluricavitaires. Il y a apparition de chambres favéolées dont les parois sont
couvertes de septa leurs conférant un aspect alvéolé. Cette structure
alvéolaire se retrouve chez les mammifères (figures 3-39,
3-40 et 3-41).
Figure 3-39: Poumons de reptiles: chez
ces espèces, l'organisation des poumons passe du système sacculaire des
batraciens au système alvéolaire des mammifères. Chez la plupart d'entre
elles, la structure générale du poumon n'est guère différente de celle du
poumon des amphibiens: il s'agit d'un poumon unicavitaire avec septa de 1er et
2ème ordre (A). Chez certaines espèces (caméléons, iguanes), 1 ou 2 septa
longitudinaux prennent de l'importance et subdivisent la cavité pulmonaire en 2
ou 3 chambres; c'est le poumon paucicavitaire (B) annonçant la structure
pluricavitaire. Chez les tortues, varans et caïmans, la structure pulmonaire se
complique par pénétration de la bronche extra-pulmonaire dans la cavité
pulmonaire qui dès lors disparaît. Cette bronche pulmonaire donne accès à
des chambres favéolées de taille extrêmement variable (C). BrE: bronche
extra-pulmonaire; BrI: bronche intra-pulmonaire; ChD: chambre dorsale; ChV:
chambre ventrale; F: favéole; S: septum. D'après Beaumont et Cassier1987,
modifié.
Figure 3-40: Poumons des mammifères. Chez les mammifères, la bronche intra-pulmonaire donnant accès aux chambres favéolées chez les reptiles, se divise régulièrement en bronchioles de divers ordres se terminant dans des sacs alvéolaires (A). Trachée, bronches et bronchioles n'ont aucun rôle respiratoire et sont couvertes d'un épithélium cilié et de cellules sécrétrices; elles conduisent l'air au système alvéolaire constitué d'un enchevêtrement de canaux alvéolaires, d'alvéoles et de capillaires sanguins (B, C, D, E). Les alvéoles communiquent de plus entre elles par des pores alvéolaires (C). A: alvéole; BrR: bronchiole respiratoire avec épithélium cilié; CA: canal alvéolaire; CM: cellule musculaire lisse; CS: capillaire sanguin; P1: pneumocyte 1 (respiratoire); P2: pneumocyte 2 (surfactant); SA: sac alvéolaire; TC: tissu conjonctif. A, C, D, E: d'après Junqueira et al. 1992, modifié. B: d'après Beaumont et Cassier 1987, modifié.
Chez les mammifères, les alvéoles se présentent comme de petits sacs d'environ 200 m de diamètre aux parois extrêmement minces, constituées essentiellement d'une fine couche de cytoplasme de pneumocytes de type 1 recouvrant une fine pellicule de cytoplasme de cellules de l'endothélium vasculaire. Cette paroi est si mince que l'on peut voir à travers la forme des globules rouges circulant dans les capillaires (figure 3-41). Cette disposition favorise évidemment les échanges gazeux qui s'opèrent à ce niveau par diffusion. La fragilité des parois des alvéoles rend indispensable la présence de surfactants, sécrétés chez tous les amniotes par les pneumocytes de type 2. La production de ces cellules aboutit à la formation d'un film monomoléculaire de nature phospholipidique qui recouvre la paroi des alvéoles. Ce film, essentiellement composé de dipalmitoyl-lécithine chez les mammifères, réduit largement la tension superficielle au niveau des alvéoles, évitant qu'elles ne collabent à l'expiration. Il réduit par ailleurs largement le travail inspiratoire en diminuant la force nécessaire à la dilatation des alvéoles.
Figure 3-41: Vues en microscopie électronique du système alvéolaire au niveau du poumon de mammifère. A: Faible grossissement (x60) montrant des bronchioles se terminant dans le système alvéolaire. B: Capillaires (C) et alvéoles (Alv) (x500). C: Alvéoles au microscope électronique à balayage, l'épaisseur des tissus entre alvéole et sang est si mince que l'on peut distinguer la forme des globules rouges dans un capillaire (x7230). A et C: d'après Junqueira et al. 1992, modifié. B: d'après Sandborn 1970, modifié.
Le système alvéolaire présente évidemment l'avantage majeur d'augmenter considérablement la surface d'échange. On estime ainsi que l'homme par exemple compterait dans ses poumons, d'un volume total moyen de quelque 4 l, plus ou moins 300 millions d'alvéoles pour une surface d'échange d'environ 85 m2. Il semble qu'il existe chez les mammifères une relation de proportionnalité directe entre la surface alvéolaire et la masse. La surface d'échange étant plus grande par unité de poids chez la musaraigne que chez l'homme par exemple (figure 3-42). Ce type de relation est à mettre en rapport avec la plus grande demande énergétique des animaux de petite taille. Elle s'explique par la présence chez ces derniers d'alvéoles de très petites dimensions, rendant la surface d'échange par unité de volume pulmonaire plus importante que chez les animaux de grande taille: 1800 cm2 par cm3 chez la musaraigne contre 550 chez le chien ou 200 chez l'homme (figure 3-43).
 |
Figure 3-42: Rapport Surface alvéolaire - Poids chez les mammifères. D'après Weibel et al. (dans Schmidt-Nielsen et al., 1980), modifié. |
Figure 3-43: Comparaison, à la même échele, de vues en microscopie électronique à balayage du parenchyme pulmonaire chez la musaraigne (A) et l'homme (B). A: alvéoles, D: canaux alvéolaires, S: septa. D'après Weibel et al. (dans Schmidt-Nielsen et al., 1980), modifié.
Un problème majeur du système réside dans l'organisation de la ventilation qui se fait en un mouvement de va-et-vient dont résulte un important volume mort qui ne participe pas aux échanges respiratoires (voir plus loin). A ce volume mort s'ajoute le volume d'air se trouvant dans les conduits ventilatoires qui lui non plus ne participe pas aux échanges au niveau alvéolaire (voir plus loin). Ces problèmes n'existent pas chez les oiseaux qui ont développé un échangeur respiratoire de type tubulaire très particulier.
b. les poumons tubulaires
Chez les oiseaux, l'échangeur respiratoire est représenté par deux poumons
rigides, inextensibles dont les unités fonctionnelles sont des parabronches
tubulaires au niveau desquels l'air passe toujours dans le même sens. La
ventilation est assurée par de volumineux sacs aériens extrapulmonaires. Il
n'existe aucune structure anatomique particulière au niveau des bronches
reliant les sacs au système parabronchique permettant de rendre compte d'un
contrôle de l'orientation du flux d'air. Il semble qu'il faille faire
intervenir un système de "valve aérodynamique" mettant en jeu la
force du courant d'air généré par les mouvements de la cage thoracique et la
résistance des bronches au passage de l'air. Dans ce cadre, des modifications
du diamètre des bronches pourraient jouer un rôle non négligeable. Ces sacs
n'interviennent pas directement dans les échanges gazeux. Ils fonctionnent
comme des soufflets assurant une circulation continue de l'air dans les
parabronches (figure 3-44).
Figure 3-44A: Poumons tubulaires des
oiseaux. Les poumons, rigides, sont organisés en un système parabronchique.
Chaque poumon est pénétré d'une bronche primaire qui se divise en de très
nombreuses bronches secondaires elles-mêmes divisées en très nombreuses
bronches tertiaires qui s'anastomosent en parallèle pour former des circuits
parabronchiques. Leur longueur varie de 1 à 4 cm et leur diamètre de 1 à 2
mm. La paroi des parabronches est percée d'innombrables orifices qui conduisent
à des chambres ou atria d'un diamètre de 1/10 mm. Les atria sont unis par un
réseau de très fins canalicules, "capillaires aériens", de 3 à 10 m, entrelacés avec
un réseau très dense de capillaires sanguins. L'air est propulsé dans les
parabronches à partir de sacs aériens antérieurs, ventraux et postérieurs.
A: système parabronchique et sacs aériens. B: Vue en microscopie à balayage
d'une section dans le système parabronchique, remarquer les entrées vers les
atria; d'après Schmidt-Nielsen 1983, modifié.

Figure 3-44B: Organisation schématique du poumon tubulaire des oiseaux; d'après Beaumont et Cassier 1987, modifié.
Ce système assure une surface d'échange pratiquement dix fois supérieure à celle obtenue chez les mammifères, à taille comparable. Il élimine par ailleurs l'essentiel du volume mort. Il assure donc un meilleur rendement par un renouvellement total de l'air au niveau des surfaces d'échange (figure 3-45). Le rendement est également amélioré par une disposition dite à "courants croisés" des flots de sang et d'air au niveau des parabronches (figure 3-46).
La paroi des parabronches est percée d'innombrables orifices qui conduisent à des chambres (atria) unies par un réseau de très fins "capillaires" aériens, canalicules d'un diamètre de 3 à 10 m, entrelacés avec un réseau de capillaires sanguins (figure 3-46). La lumière des capillaires aériens résulterait de cavités intracellulaires qui fusionneraient entre elles au niveau de pneumocytes de type 1. Cette disposition amènerait l'épaisseur totale de la barrière à la diffusion à 0,1 ou 0,2 m, soit nettement moins que chez les mammifères (1 à 5 m). Les pneumocytes de type 2 sont situés en dehors de ce système, à la surface des atria et des parabronches.
 |
|
3.4.2. Le système trachéen des insectes
Nous avons déjà brièvement introduit quelques notions concernant ce système
en parlant des insectes aquatiques (cfr. 3.2.3.b).
Les trachées sont des invaginations ectodermiques extrêmement ramifiées qui s'étendent jusqu'au niveau cellulaire. Chez les insectes, l'air est donc amené de trachées en trachéoles directement aux cellules où s'effectuent les échanges gazeux (figure 3-47). Le sang n'a donc chez ces espèces aucune fonction respiratoire, à l'encontre de ce qui se passe chez toutes les autres espèces. Dans ce cadre, il est dépourvu de pigments transporteurs d'oxygène.
Figure 3-47: Le système respiratoire des insectes. A: Trachée et trachéoles amenant l'air directement au niveau des tissus où les échanges gazeux s'effectuent directement, sans passage par le système circulatoire. D'après Eckert et Randall 1983, modifié. B: Vue en microscopie électronique d'un muscle de cycadide montrant les trachéoles du système respiratoire (Tr) entre les fibres musculaires (M). D'après Schmidt-Nielsen 1983, modifié.
Les trachées débouchent à l'extérieur par des stigmates: orifices pairs, symétriques, à disposition primitivement segmentaire. Le nombre de paires est très variable chez les insectes terrestres adultes. Dans la plupart des cas, il est de 10 avec deux paires thoraciques et huit abdominales. Chez les insectes dits holopneustiques, tous les stigmates sont ouverts et fonctionnels. Quelques-uns seulement restent en fonction chez les insectes dits hemipneustiques. Les stigmates communiquent avec de larges troncs trachéens longitudinaux qui envoient de nombreuses ramifications vers tous les organes (figure 3-48). Les variations sur ce modèle de base sont nombreuses et leur étude sort du cadre de la physiologie. Notons simplement que chez de nombreux insectes bons voiliers (hanneton, abeille, mouche par exemple), le système trachéen est en relation avec des sacs aériens servant de réservoirs. Ils peuvent être aisément comprimés par les mouvements ventilatoires, ce qui parait améliorer la ventilation au niveau tissulaire. Chez le hanneton, ces sacs sont petits mais très nombreux; le volume d'air emmagasiné à leur niveau représente près de la moitié du volume de l'insecte. Chez les abeilles et les mouches, les sacs sont peu nombreux et très grands; ils occupent l'essentiel de la cavité abdominale. Le volume d'air emmagasiné peut représenter ici largement plus de la moitié du volume total de l'insecte.
 |
Figure 3-48: Le système trachéen des insectes. A: Système simple avec trachées anastomosées. B: Système avec sacs aériens. D'après Beaumont et Cassier 1983, modifié. |
3.4.3. Conduits ventilatoires, échanges
hydriques et thermiques
Pour garder toute son efficacité dans les échanges gazeux, le fragile
épithélium respiratoire doit être maintenu constamment humide. Les poumons
représentent donc une importante surface humide en contact avec un air
relativement plus sec venant de l’extérieur et, dès lors, sujette à l'évaporation. L'air au
niveau alvéolaire est constamment maintenu à saturation d'humidité malgré
une entrée d'air extérieur sec grâce essentiellement au système d'échangeur de chaleur et
d'humidité que constituent les conduits ventilatoires d'une part et grâce aussi à
l'important volume d'air ne participant pas directement aux échanges d'autre
part. En fait, lors de l'inhalation, l'air extérieur passe dans une série de
conduits respiratoires au niveau desquels il va pouvoir se réchauffer et
s'hydrater pour atteindre la saturation en humidité et la température requise
au niveau alvéolaire. Chez les mammifères par exemple, l'air au niveau
alvéolaire est à 37 °C et saturé en eau. Considérons un air entrant à 15
°C et à 20 % d'humidité relative. Cet air va gagner de la chaleur et de la
vapeur d'eau au fur et à mesure qu'il avance dans les conduits nasaux, les
bronches, etc... pour atteindre 37 °C et la saturation au niveau de la surface
d'échange (figure 3-49). Les conduits cédant de la
chaleur vont dans le même temps se refroidir. A l'expiration, l'air saturé et
à 37 °C va perdre de la chaleur au niveau des conduits. Cette baisse de
température va provoquer une condensation d'eau qui sera récupérée et
servira à l'hydratation de l'air frais entrant. L'air sortant sera saturé en
humidité et à une température intermédiaire entre celle de l'air extérieur
(15 °C) et celle de l'air au niveau pulmonaire (37 °C). Ce système permet
donc le maintien de l'humidité nécessaire au niveau pulmonaire en limitant les
pertes d'eau. Nombre d'animaux désertiques pour lesquels l'économie de l'eau
est importante ont, en général, des conduits nasaux assez longs, de façon à
récupérer un maximum d'eau de l'air sortant des poumons. Dans certains cas, ce
système est d'importance critique dans la balance hydrique et la survie des
animaux (voir chapitre 1). Ainsi par exemple, le
rat kangourou (Dipodomys spectabilis) pourra vivre sans problème
dans le désert, sans rien boire, en se nourrissant uniquement d'éléments
végétaux secs et en respirant un air à 25 °C et à 20 % d'humidité
relative. Dans un air plus sec (10 % H.R.), à la même température, il est
incapable de maintenir une balance hydrique correcte et meurt rapidement.
Figure 3-49: Echanges de chaleur et d'eau au niveau des conduits respiratoires chez un mammifère. HR: humidité relative. *: la température est donnée à titre indicatif. Elle peut varier en fonction de la longueur des conduits et de leur capacité d'échange de chaleur, du volume d'air exhalé et de sa vitesse de sortie.
La quantité d'eau récupérée dans le système d'échange de chaleur sera évidemment fonction de la température extérieure et de son degré d'humidité relative. Le maintien de l'air à saturation en eau au niveau alvéolaire va évidemment être fonction des mêmes paramètres mais aussi du volume d'air effectivement renouvelé à ce niveau. Dans ce cadre il est intéressant de remarquer que la fraction d'air renouvelée au niveau alvéolaire à chaque inspiration ne représente en conditions normales qu'une petite partie du volume total d'air contenu dans l'ensemble du système respiratoire. Chez l'homme par exemple, sur un volume total possible d'air d'environ 5 litres, quelque 350 à 600 ml d'air seulement seront remplacés au niveau alvéolaire à chaque inspiration (voir 3.4.4 ci-après).
3.4.4. La ventilation et son contrôle
a. mouvements et mécaniques ventilatoires
La ventilation des cavités pulmonaires et son contrôle n'ont été que peu
étudiés dans la plupart des cas.
Il semble que chez bon nombre d'invertébrés terrestres, la ventilation n'est pas organisée de façon spécifique, les variations de volume des cavités pulmonaires sont essentiellement en relation avec les mouvements du corps.
Chez les insectes, les échanges respiratoires sont également passifs chez les espèces de petite taille au métabolisme peu élevé. La circulation de l'air dans les trachées résulte d'un simple phénomène de diffusion. En revanche, les insectes aériens de grande taille ou les espèces actives effectuent des mouvements ventilatoires spécifiques auxquels participent les muscles abdominaux, dorso-ventraux et longitudinaux. La contraction des muscles diminue en fait le volume de la cavité viscérale et l'augmentation de pression interne résultante chasse l'air de l'appareil trachéen. Chez ces espèces, c'est donc l'expiration qui est active. L'inspiration est un phénomène passif, les parois du corps revenant à leurs positions initiales par le seul effet de l'élasticité lorsque les muscles se relâchent. En fonction des muscles mis en jeu, la circulation de l'air peut ainsi être mise en rapport essentiellement avec un aplatissement dorso-ventral de l'abdomen (carabes, criquets), des contractions télescopiques des segments abdominaux (mouches, abeilles, dytiques, hydrophiles) ou encore des mouvements de type péristaltique de l'abdomen (tipules). Beaucoup d'insectes peuvent pratiquer une ventilation dite discontinue. Dans ce système, les spiracles restent fermés pendant un certain temps, pouvant varier de quelques minutes à plusieurs heures suivant l'espèce et le niveau d'activité de l'animal. Pendant cette phase, le CO2 s'accumule dans les trachées alors que la teneur en oxygène diminue largement. Cette phase est suivie d'une phase de "fluter", en général courte, pendant laquelle les spiracles s'ouvrent et se ferment très rapidement. Il y a ensuite une phase d'ouverture pendant laquelle l'essentiel des échanges gazeux s'effectue. A l'heure actuelle, la finalité de ce système n'est pas totalement claire. Décrit au départ chez des larves d'insectes vivants dans des milieux très secs, on avait imaginé qu'il permettait une certaine économie d'eau en limitant les échanges gazeux et donc l'évaporation. Le fait que beaucoup d'insectes, même d'habitats humides, pratiquent la ventilation discontinue met à l'heure actuelle cette hypothèse directement en question.
Tous les vertébrés pulmonés ont des systèmes de ventilation plus ou moins efficace. Les appareils ventilatoires spécifiques n'apparaissent toutefois qu'à partir des reptiles. Chez les poissons pulmonés, l'inspiration est assurée par compression du plancher buccal qui force l'ouverture de la glotte. L'expiration est essentiellement passive. La contraction de muscles lisses intrinsèques au système respiratoire peut toutefois y participer de manière plus ou moins importante. Chez les batraciens, le système est assez proche. Chez la grenouille par exemple, l'air est inhalé dans la cavité buccale par abaissement du plancher de celle-ci. L'air vicié se trouvant dans les poumons est ensuite expiré par contraction musculaire et passe au-dessus de l'air frais accumulé dans la bouche. Les narines sont ensuite fermées et l'air est poussé vers les poumons lorsque le plancher buccal se soulève pour reprendre sa position de départ, ce qui permet au cycle de recommencer (figure 3-50).
Figure 3-50: Le cycle respiratoire chez la grenouille - explication dans le texte. D'après Gans et al. (Science 163, 1223, 1969), modifié.
Chez les reptiles, les oiseaux et les mammifères, la ventilation met en jeu les mouvements d'une "cage thoracique" organisée de différentes façons suivant les groupes (figure 3-51). Le système fonctionne en général comme une pompe aspirante, parfois aspirante - foulante mais jamais uniquement foulante. Le renouvellement de l'air au niveau de l'épithélium respiratoire est assuré par des modifications de volume de la cage obtenues par contraction de muscles striés et relaxation avec retour à une position de repos. L'inspiration est toujours liée à une phase active impliquant la contraction de muscles particuliers. L'expiration sera en général largement passive, à mettre en rapport avec un retour des tissus plus ou moins étirés à l'inspiration à une position de repos. Dans de nombreux cas, l'expiration, de passive en conditions normales, peut devenir active, mettant en œuvre la contraction de muscles spécifiques. Inspiration comme expiration peuvent être aidées par des mouvements du corps. Ainsi, lors de la course chez les quadrupèdes lorsque les pattes de devant touchent le sol, la cage thoracique peut être comprimée par les forces de réactions du sol à travers les membres antérieurs et par un mouvement en avant des viscères. Dans de nombreux cas, le rythme respiratoire devient ainsi couplé au rythme de la course; la respiration du cheval au galop est un bel exemple de ce type de synchronisation (figure 3-52). Chez les oiseaux en vol, la respiration peut être rythmée par les coups d'aile. Les muscles des ailes sont attachés à un sternum très mobile qui ferme la cage thoracique ventralement. Un mouvement d'aile vers le bas fait monter le sternum et comprime la cage thoracique; un mouvement vers le haut à l'effet inverse (figure 3-51). Ce type de synchronisation se remarque également chez certains insectes où la ventilation augmente avec le rythme des battements d’ailes au cours du vol (Komai, J. Exp. Biol., 201, 2359, 1998). Ces modifications sont dues à des variations du volume des trachées en rapport avec les mouvements de compression de la région thoracique qu’implique le mouvement des ailes chez certains insectes (voir chapitre 7).
Figure 3-51: Les mouvements
respiratoires chez les vertébrés pulmonés à partir des reptiles. L'air entre
et sort des poumons en fonction des mouvements d'une cage thoracique à laquelle
ils sont solidarisés par l'intermédiaire d'un système de plèvres. La cage
est fermée par des côtes attachées dorsalement à la colonne vertébrale et
ventralement à un sternum. L'inspiration est toujours active: la contraction
des intercostaux externes tire les côtes vers l'extérieur, mettant les poumons
en dépression, ce qui provoque l'entrée d'air. L'expiration peut être active,
impliquant la contraction d'intercostaux internes. Elle peut aussi être
passive, dépendant de l'élasticité des tissus étirés à l'inspiration. Ce
système de base est mis en œuvre tel quel chez les lézards (A). Chez les
oiseaux, le sternum est très mobile et sert de point d'attache aux muscles du
vol; ses mouvements sont ainsi associés aux mouvements des côtes mais aussi
des ailes (B). Chez les crocodiliens et les mammifères, un diaphragme
musculaire ferme la cage thoracique à l'arrière, formant une paroi bombée; la
contraction aplatit le diaphragme, assistant l'inspiration de façon essentielle
chez les mammifères, moindre chez les crocodiles (C et D). L'expiration peut
être plus ou moins active, impliquant la contraction des intercostaux internes
mais aussi des abdominaux. Ce dernier processus est important chez les
crocodiles où il intervient dans la respiration normale; le foie notamment est
ici poussé vers l'avant et comprime les poumons à chaque expiration (C). Les
serpents ont un ou deux poumons très longs présentant une partie antérieure
respiratoire accolée à la paroi dorsale et une partie postérieure non
respiratoire servant de sac aérien. Les côtes, présentes sur toute la
longueur provoquent un flux inspiratoire qui peut être limité à une seule
région lorsque l'animal avale une proie. Lorsque la proie est dans la partie
antérieure, la ventilation est obtenue par dilatation de la partie postérieure
sacculaire. L'air passe alors sur les favéoles situées dorsalement au-dessus
de la proie (E). Chez les tortues, la cage thoracique est soudée à la
carapace. Les plèvres entourant les poumons sont ici étirées vers l'avant et
l'arrière par des faisceaux de muscles particuliers attachés à la carapace
(F). G: Intercostaux et mouvements des côtes chez les mammifères. A à G:
d'après Kardong 1995, modifié.
Figure 3-52: Synchronisation de la respiration avec le galop chez le cheval. L'enregistrement des bruits de l'air au niveau des nasaux indique les phases d'inspiration (I) et d'expiration (E). L'inspiration commence lorsque le cheval s'étend, au début d'un bond en prenant appui sur ses jambes postérieures (1). L'expiration se termine lorsque le cheval termine le bond, en se recevant sur les antérieures (3). D'après Kardong 1995, modifié.
b. cycles et fractions respiratoires
Dans la plupart des cas, le renouvellement de l'air au niveau des surfaces
d'échange est effectué au cours d'un seul cycle inspiration-expiration. Chez
les oiseaux cependant, la disposition très particulière de l'échangeur par
rapport aux réserves d'air des sacs aériens fait qu'il faut deux cycles pour
que tout l'air inhalé lors d'une première inspiration passe sur le système
parabronchique (figure 3-53). De plus chez ces espèces,
tout l'air inhalé participe aux échanges gazeux. C'est loin d'être le cas
dans les autres groupes chez lesquels la respiration met en jeu un va et vient
de l'air dans les structures respiratoires, qui génère, comme nous l'avons
déjà signalé, un volume "mort" ou "résiduel" d'air qui
ne participe pas aux échanges.
Figure 3-53: Mouvement de l'air inhalé lors d'une première inspiration au cours de deux cycles respiratoires chez l'autruche. D'après Schmidt-Nielsen 1983, modifié.
Si le volume total d'air inspiré peut être éminemment variable en fonction des conditions, le volume mort lui peut être considéré comme pratiquement invariable, contenu dans des structures (essentiellement conduits respiratoires mais aussi alvéoles ne participant pas aux échanges) au volume total fixe ou, en tout cas, très largement moins extensibles que les poumons. La capacité totale du système respiratoire sera donc égale à l'ensemble du volume mort et du volume d'air maximum qui peut être inspiré et expiré. Ce dernier volume correspond à ce qu'il est convenu d'appeler la capacité vitale. Celle-ci peut se décomposer en un volume courant, un volume inspiratoire de réserve et un volume expiratoire de réserve (figure 3-54). Le volume courant correspond au volume d'air effectivement renouvelé lors d'un cycle respiratoire en conditions de repos. Les volumes de réserve sont ceux qui peuvent être renouvelés au cours d'inspirations ou d'expirations plus ou moins profondes. Le volume expiratoire de réserve additionné du volume mort constitue en fait le volume minimum se trouvant dans le système respiratoire au repos avant inspiration. C'est ce qu'il est convenu d'appeler la capacité résiduelle fonctionnelle du système. Son volume, considéré par rapport au volume courant, intervient de façon essentielle dans l'efficacité de l'échangeur respiratoire. En effet, plus ce volume sera grand par rapport au volume courant, moins il y aura d'air effectivement renouvelé au niveau alvéolaire. Ceci signifie que l'air entrant sera mélangé avec un volume d'air stagnant d'autant plus grand et, dès lors, la pression partielle de l'oxygène au niveau alvéolaire sera moindre alors que la pression partielle en CO2 sera plus élevée. Ainsi, chez l'homme au repos, la capacité résiduelle fonctionnelle est d'environ 2 l et le volume courant de 0,6 l. Ceci signifie qu'au cours d'une inspiration, 600 ml d'air frais seulement entrent dans un système contenant déjà 2 l d'air. Le renouvellement ne porte donc dans ces conditions que sur quelque 30 % de l'air accessible pour l'échange gazeux (contre quelque 90 % pour les poumons tubulaires des oiseaux). Il en résulte que l'air alvéolaire est de composition pratiquement constante au repos avec une pression partielle en O2 et CO2 de plus ou moins 100 et 40 mmHg alors que les pressions sont respectivement de 150 et 0,2 mmHg dans l'air extérieur.
Figure 3-54: Volumes et capacités respiratoires chez l'homme et par extension chez les autres pulmonés.
Toute variation du volume courant et/ou du volume résiduel fonctionnel va modifier les pressions partielles en O2 et en CO2 et donc l'efficacité de l'échange respiratoire. Ainsi, l'utilisation chez l'homme d'un tube donnant accès à l'air extérieur pour respirer (plongée, avalanche, effondrement) augmente en fait le volume mort et donc la capacité résiduelle fonctionnelle avec pour conséquence néfaste une augmentation du CO2 et une diminution de l'O2 au niveau alvéolaire. Comme nous le verrons dans la section suivante, ces modifications sont compensées par des variations de volume tidal et de fréquence respiratoire. A la limite, la respiration devient impossible à partir d'une certaine longueur de tube. Différents animaux au long cou tels que le cygne (figure 3-55) ou la girafe ont ainsi un volume mort relativement important. Celui-ci est en général compensé par un volume tidal par rapport à la capacité pulmonaire totale plus important que la moyenne.
Il est par ailleurs clair que tous les paramètres pouvant modifier l'ampleur des mouvements respiratoires tels que la pression en plongée (figure 3-65) ou la diminution d'élasticité pulmonaire observée dans l'emphysème par exemple vont aussi aboutir à une insuffisance respiratoire due à une augmentation de la capacité résiduelle fonctionnelle à laquelle s'ajoute une diminution concomitante du volume courant.
Dans l'exercice, il y a également variation concomitante du volume tidal et de la capacité résiduelle fonctionnelle. Dans ce cas, à l'inverse de ce qui se passe dans l'insuffisance respiratoire, le volume tidal augmente et la capacité résiduelle fonctionnelle diminue. Ceci aura pour conséquence une meilleure ventilation pulmonaire. On n'observe cependant pas de variations significatives de la PO2 et de la PCO2 au niveau alvéolaire. Ces variations sont en effet contrebalancées par l'augmentation de demande en O2 et de rejet en CO2 qu'engendrent l'augmentation d'activité métabolique. Ceci nous amène à envisager le contrôle de la respiration, notamment en fonction des nécessités métaboliques.
c. régulation de la respiration
En général, l'activité des systèmes respiratoires peut être ajustée en
fonction de deux nécessités premières: pourvoir l'organisme de la quantité
en oxygène dont il a besoin et éliminer le CO2 résultant de ces
activités métaboliques. De très nombreux travaux montrent dans ce cadre qu'un
contrôle de la ventilation pulmonaire en fonction de la pression partielle en O2
du sang existe dans tous les groupes zoologiques. Un contrôle en fonction du pH
sanguin et de sa pression partielle en CO2 parait également assez
général, du moins chez les vertébrés. La sensibilité de ce dernier système
est cependant éminemment variable, étant essentiellement développée chez les
mammifères. Un contrôle en fonction des teneurs en O2 et CO2
du milieu extérieur pourrait également exister chez certaines espèces
amphibies.
La régulation de l'activité respiratoire n'a cependant été étudiée de façon détaillée que chez très peu d'espèces, essentiellement des mammifères, rendant une approche comparée difficile. Nous nous concentrerons donc sur le modèle mammalien.
L'activité respiratoire s'effectue sur un rythme de base qui peut être augmenté ou diminué, cette régulation visant en fin de compte au maintien des PO2 et PCO2 sanguines par des ajustements du débit ventilatoire au niveau alvéolaire. Une diminution de PO2 et/ou une augmentation de PCO2 vont induire une augmentation de l'activité respiratoire qui se traduira par une hyperventilation (augmentation du débit ventilatoire) au niveau alvéolaire. A l'inverse, une augmentation de PO2 et/ou une diminution de PCO2 induiront une diminution d'activité respiratoire et une hypoventilation alvéolaire. Les modifications de ventilation seront donc essentiellement déterminées par, et tendront à corriger, des états d'hypoxie ou d'hyperoxie ou encore d'hypo ou d'hypercapnie sanguine (voir plus loin, 3 et figure 3-58).
Le débit ventilatoire représente en fait la quantité d'air renouvelé par unité de temps (l/min en général). Il est donc fonction de la fréquence des cycles respiratoires inspiration-expiration et du volume courant, c'est-à-dire de la quantité d'air inspirée par cycle.
Les modifications de ventilation sont toujours associées à une variation de la fréquence des cycles. Dans de nombreux cas, une variation de l'ampleur de la respiration (variation du volume courant) intervient également. Ainsi, chez l'homme au repos, la fréquence respiratoire moyenne est d'environ 12 inspirations/minutes et le volume courant de 0,7 l; ces valeurs passent respectivement à 20 et 2 lors d'un exercice soutenu et peuvent atteindre 40 et 3 en travail maximum. Le débit ventilatoire pourra ainsi passer de 7 l à plus de 100 l !
Le contrôle de l'activité respiratoire s'effectue essentiellement par voie nerveuse, impliquant des centres de commandes bulbaires recevant des afférences centrales et périphériques et ayant des efférences contrôlant l'activité des muscles inspirateurs et expirateurs (nerfs phréniques et intercostaux).
1. Les centres respiratoires
Les études actuelles ne permettent pas encore de comprendre parfaitement
l'organisation du contrôle de l'activité ventilatoire cyclique. Quatre centres
bulbaires sont classiquement impliqués chez l'homme: un centre apneustique, un
centre pneumotaxique, un centre inspiratoire et un centre expiratoire (figure 3-56).
L'excitation du centre expiratoire entraîne un arrêt de l'activité inspiratoire et une contraction des muscles expiratoires. A l'inverse, l'excitation du centre inspiratoire provoque une contraction des muscles inspiratoires (diaphragme et intercostaux externes - cfr. figure 3-51) ainsi qu'une relaxation des muscles expiratoires (intercostaux internes). L'activité du centre inspiratoire serait contrôlée par le centre apneustique, lui-même contrôlé par le centre pneumotaxique (figure 3-56). Le centre apneustique stimulerait en permanence le centre inspiratoire et son activité serait inhibée de façon rythmique par le centre pneumotaxique.
 |
|
L'activité de ces centres serait contrôlée par voie centrale et périphérique.
Les influences centrales restent mal étudiées. De nombreuses observations indiquent cependant que des mécanismes nerveux centraux peuvent participer à l'adaptation du débit ventilatoire. Ainsi, on peut exercer un contrôle volontaire sur la respiration (apnée en plongée notamment). Une sensation douloureuse ou l'application soudaine d'eau froide sur la peau vont s'accompagner d'une hyperventilation. De même, une flexion-extension volontaire des jambes provoque une hyperventilation immédiate, un phénomène qui est d'intérêt pour les athlètes qui vont entamer une compétition. Il semble par ailleurs que dans l'émission de sons, les modulations d'activité respiratoire soient obtenues par effet direct de centres corticaux sur les neurones moteurs commandant les muscles respiratoires, les centres respiratoires bulbaires n'interviendraient pas.
Les influences périphériques sont essentiellement fonction de conditions physico-chimiques locales (O2, CO2, pH sanguin, etc...) et d'informations mécaniques provenant des poumons, ces dernières intervenant plus particulièrement dans le contrôle de la rythmicité ventilatoire.
2. Les stimuli mécaniques et la rythmicité
La perception de différents stimuli mécaniques est à la base de différents
réflexes qui vont régler la rythmicité de la respiration.
1) Réflexe inhibiteur de l'inspiration
La distension des poumons à l'inspiration est perçue par des
mécanorécepteurs situés sur les parois des bronches terminales. Leur
excitation va provoquer une inhibition de l'inspiration apparemment via
inhibition du centre apneustique. Ce mécanisme est appelé réflexe de
distension pulmonaire ou réflexe de Brewer-Hering, du nom des scientifiques qui
l'ont abordé chez l'homme pour la première fois en 1868.
2) Réflexe excitateur de l'inspiration
La diminution de volume pulmonaire à l'expiration induit une contraction du
diaphragme et donc l'inspiration. La nature des récepteurs sollicités reste
inconnue. Ce réflexe ferait intervenir le centre apneustique et le centre
inspiratoire.
3) Réflexe proprioceptif des muscles respiratoires
L'étirement des fibres musculaires, essentiellement au niveau des intercostaux
et à un degré moindre du diaphragme est perçue par des mécanorécepteurs de
type annulo-spiraux (figure 3-57). Ceux-ci vont induire par
voie réflexe une contraction du muscle sur lequel ils se trouvent. Il s'agit
d'un réflexe spinal de type myotatique (cfr. relations
nerveuses); les centres respiratoires n'interviennent donc pas.
Figure 3-57: Réflexe myotatique spinal et rythmicité respiratoire. Dans les muscles intercostaux et le diaphragme se trouvent des fuseaux neuromusculaires contenant des récepteurs annulo-spiraux sensibles à l'allongement. Leur stimulation provoque par voie réflexe spinale une contraction des muscles sur lesquels ils se trouvent. Ainsi l'inspiration provoque un étirement des muscles intercostaux internes, ce qui provoque leur contraction. L'allongement du fuseau peut également être obtenu par contraction des fibres musculaires auxquelles il s'attache sous l'action de neurones moteurs g. CA et CP: cornes antérieur et postérieure de la moelle, IA: fibre sensitive. D'après Flandrois et al. 1976, modifié.
3. Les stimuli chimiques et l'adaptation de
la ventilation
Comme nous l'avons déjà signalé, les variations de PO2, de PCO2
et de pH au niveau sanguin provoquent rapidement des modifications ventilatoires.
Les variations de ces paramètres sont perçues au niveau de chémorécepteurs
qui agissent d'une façon encore peu claire sur les centres respiratoires
bulbaires.
Les récepteurs sensibles à la tension en oxygène sont logés dans la crosse aortique ainsi que dans les corpuscules carotidiens, à la bifurcation des artères carotides. Les récepteurs sensibles au pH et au CO2 se trouvent au niveau du bulbe céphalo-rachidien, en connexion pratiquement directe avec les centres respiratoires.
Une hypoxie (PO2 ¯) ainsi qu'une hypercapnie (PCO2 ) ou une baisse de pH induisent une hyperventilation corrective (figure 3-58). Ces réactions peuvent parfaitement rendre compte de l'augmentation de débit ventilatoire observée au cours d'un exercice par exemple (figure 3-59). A l'inverse, une hyperoxie (PO2 ), une hypocapnie (PCO2 ¯) ou une augmentation de pH provoquent une hypoventilation (figure 3-58). La réponse à l'hypercapnie (PCO2 ) n'est bien marquée que chez les mammifères. Elle est peu marquée voir peu significative chez les invertébrés. Elle reste peu importante chez les vertébrés amphibies pouvant accumuler du CO2 en milieu aérien (poissons pulmonés, batraciens). Elle est également moindre chez les oiseaux que chez les mammifères. Le peu de sensibilité relative des oiseaux à la PCO2 les aident vraisemblablement à supporter des variations importantes et brusques de l'altitude (voir chapitre 4, 3). Chez les mammifères eux-mêmes, on note de larges variations dans l'importance de la réponse à l'hypercapnie. Elle est par exemple nettement moindre chez les espèces vivant dans des terriers ou chez celles pratiquant la plongée, deux conditions favorisant l'accumulation de CO2.
Figure 3-58: Réponse ventilatoire à une variation de PO2 ou de PCO2 chez l'homme. L'hyperoxie et l'hypercapnie sont obtenues par inhalation d'un mélange gazeux soit sans CO2 soit avec 6,7 % de CO2 au lieu de 5. L'hypoxie est obtenue par inhalation d'azote et l'hypocapnie par un profond soupir volontaire. D'après Flandrois et al. 1976, modifié.
La plupart du temps, les stimuli CO2 et pH sont liés, une variation de teneur en CO2 sanguine entraînant une variation de pH (voir chapitre 4). Les récepteurs impliqués sont cependant différents et il est possible de dissocier leurs effets au niveau ventilatoire (figure 3-60).
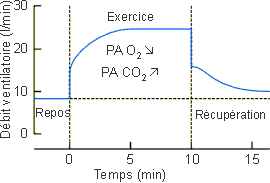 |
Figure 3-59: Evolution du débit ventilatoire au cours de l'exercice. Représentation semi-schématique. D'après Flandrois et al. 1976, modifié. |
4. Autres stimuli
Différents autres stimuli induisent des modifications intéressantes de la
ventilation. Retenons par exemple qu'une augmentation des catécholamines
(adrénaline, noradrénaline) circulantes provoque une hyperventilation. Comme
nous le verrons par la suite il s'agit là d'une réaction intéressante en cas
de stress et aussi au cours de l'exercice, conditions dans lesquelles ces
hormones augmentent dans le sang. A l'inverse de cette réaction, une
augmentation très importante d'adrénaline peut provoquer une apnée; le
blocage respiratoire qu'entraîne parfois une émotion violente est connu depuis
longtemps.
Une hyperventilation est par ailleurs classiquement observée en cas d'hypotension hémorragique grave ou lors d'une élévation de température corporelle. Chez certaines espèces utilisant le halètement pour se refroidir (voir chapitre 10), l'augmentation de température induit une polypnée importante caractérisée par une forte augmentation de la fréquence respiratoire et une diminution du volume courant. Ces variations sont favorables à une augmentation de ventilation des espaces morts, essentiellement des voies aériennes supérieures qui interviennent dans les échanges thermiques entre l'air respiré et l'organisme (cfr. 3.4.3 ci avant).