De nombreuses espèces à respiration aérienne pratiquent largement la plongée. Chez les invertébrés, c'est le cas notamment du dytique et d'autres insectes plongeurs. Ces espèces partent avec une réserve d'air extérieur qui sert de branchie physique. Dans les meilleurs des cas, les échanges gazeux s'effectuant entre l'eau et la réserve d'air sont suffisants pour assurer les échanges de l'animal qui n'a dès lors pratiquement plus besoin de faire surface bien que restant à respiration typiquement aérienne. Nous avons envisagé ce système précédemment, en considérant la respiration en milieu aquatique (cfr. 3.2.3.b, ci avant). Chez les vertébrés, on trouve des animaux plongeurs dans tous les groupes, des batraciens aux mammifères. En plongée, ces animaux sont privés d'accès à l'oxygène et un état hypoxique puis anoxique devrait s'installer rapidement. Ils peuvent cependant rester actifs sous l'eau pendant des temps relativement longs, plus longs en tout cas que ne pourraient le laisser supposer leurs possibilités thoraciques de stockage de l'oxygène. Il faut noter ici que certains reptiles comme le serpent marin Pelamis platurus et surtout les batraciens sont largement aidés par les possibilités qu'ils ont d'effectuer une respiration cutanée. Ce n'est guère le cas pour de nombreux autres reptiles (tortues, iguanes, crocodiles) ni pour les oiseaux ou les mammifères plongeurs.
Ces animaux vont en fait avoir à résoudre deux problèmes en rapport avec la plongée. Le premier est de rester actif un maximum de temps avec au départ une réserve d'oxygène limitée. Il concerne en fait l'économie d'oxygène. Le second est d'éviter les embolies gazeuses (aéroembolisme - mal des caissons) que pourraient provoquer les remontées trop rapides suite à des plongées de longue durée à grande profondeur. L'homme, mal adapté à la plongée, doit encore faire face à d'autres problèmes que nous envisagerons brièvement plus loin.
Ces deux problèmes majeurs ont été solutionnés de façon similaire dans les différents groupes de vertébrés plongeurs. Nous envisagerons les solutions adoptées en prenant l'exemple du phoque.
Les adaptations particulières en relation avec ce problème qui peuvent être mises en évidence chez le phoque concernent 1) une modification des possibilités de stockage de l'oxygène; 2) un contrôle du flot sanguin de façon à privilégier l'oxygénation des tissus ne supportant guère l'hypoxie ou l'anoxie.
Le tableau 3-6 donne une idée des possibilités de stockage de l'oxygène chez l'homme et chez le phoque.
TABLEAU 3-6
Stockage de l'oxygène chez l'homme et le phoque
| O2 (ml) | O2 (ml/kg) | % du total | |
| HOMME (70 kg) | |||
| Poumons (4,5 l, 16% O2) | 720 | 10,3 | 33,5 |
| Sang (5 l, 200 ml O2/l) | 1000 | 14,3 | 46,0 |
| Muscle (16 kg, 15 ml O2/kg) | 240 | 3,4 | 11,5 |
| Eau tissulaire (40 l, 5 ml O2/l) | 200 | 2,8 | 9,0 |
| Total | 2160 | 30,8 | |
| PHOQUE (30 kg) | |||
| Poumons (350 ml, 16% O2) | 55 | 1,8 | 3,5 |
| Sang (4,5 l, 250 ml O2/l) | 1125 | 37,5 | 72,5 |
| Muscle (6 kg, 45 ml O2/kg) | 270 | 9,0 | 17,5 |
| Eau tissulaire (20 l, 5 ml O2/l) | 100 | 3,3 | 6,5 |
| Total | 1550 | 51,6 |
Comme on le voit, l'homme possède un volume pulmonaire largement supérieur au phoque. On y retrouvera quelque 33 % de l'oxygène total stocké. Chez le phoque les poumons ne contiennent guère que 3,5 % de l'oxygène total. Le volume sanguin du phoque est par contre proportionnellement beaucoup plus important que celui de l'homme. Sa capacité à fixer l'oxygène est également supérieure. Il en résulte que 72 % de l'oxygène total sont stockés à ce niveau chez le phoque contre 46 % chez l'homme. Le tissu musculaire du phoque montre également une capacité de stockage d'oxygène 3 fois supérieure à celle de l'homme. En bref, il s'avère que 1) l'essentiel de l'oxygène se trouve stocké sous forme dissoute dans le sang et les tissus du phoque alors qu'une quantité non négligeable d'oxygène se trouve sous forme gazeuse dans les poumons chez l'homme; 2) Le phoque arrive à stocker 1,7 fois plus d'oxygène par unité de poids que l'homme. Cette quantité d'oxygène, bien que plus large que celle stockée chez l'homme, ne pourrait cependant pas permettre au phoque de tenir plus de 5 minutes en plongée en considérant une dépense énergétique normale de repos. Or l'animal peut être très actif sous l'eau puisqu'il y cherche sa nourriture, et le temps de plongée peut aller jusqu'à 15 minutes et plus. En fait le phoque en plongée modifie considérablement l'irrigation de ses masses tissulaires ce qui lui permet une économie d'oxygène. Ce point est illustré dans la figure 3-61 qui montre que seule l'irrigation du système cérébral est maintenue à une valeur constante. Tous les autres tissus subissent une diminution d'irrigation, celle-ci étant particulièrement importante au niveau des masses musculaires et splanchniques. La circulation à ces niveaux est en fait pratiquement arrêtée. Ceci présente le double intérêt de 1) laisser l'oxygène disponible pour le tissu nerveux qui supporte très mal l'hypoxie, 2) empêcher le CO2 qui s'accumule au niveau des masses musculaires d'augmenter trop rapidement dans le sang et de retarder dès lors les réflexes de remontée et d'hyperventilation qu'induit normalement l'augmentation de tension en CO2 du sang via des chémorécepteurs vasculaires (voir plus loin). Il semble par ailleurs que chez les mammifères plongeurs en tout cas, des récepteurs situés au niveau de la bouche, du nez et/ou de la glotte (en fonction de l'espèce) détectent la présence de l'eau induisant une inhibition du réflexe inspiratoire. Les informations venant de ces récepteurs seraient en fait antagonistes de celles venant des chémorécepteurs vasculaires.
Figure 3-61: Modifications de l'irrigation au niveau de différents organes d'un phoque de Weddell pendant la plongée.
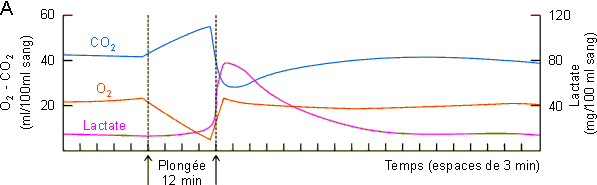
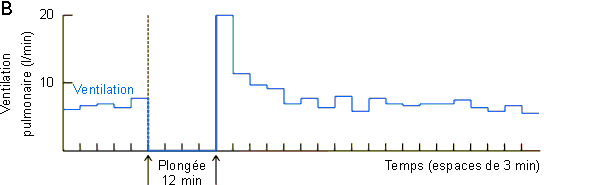
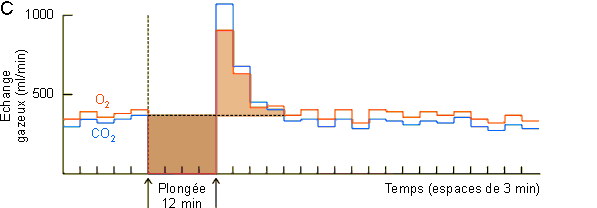
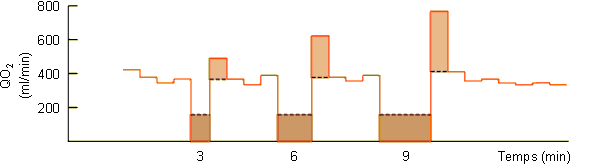
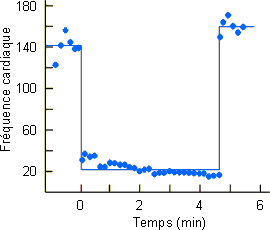
Ces modifications du flot sanguin déterminent par ailleurs la mise en route d'un métabolisme anaérobie avec production de lactate. Comme le montre la figure 3-62A une augmentation de lactate ne se marquera vraiment au niveau sanguin qu'en fin de plongée, lors du rétablissement de la circulation au niveau des masses musculaires et splanchniques. La mise en circulation de l'acide lactique et du CO2 accumulés pendant la plongée va provoquer via des chémorécepteurs vasculaires une hyperventilation (figure 3-62B) favorable dans le contexte d'augmentation de consommation d'oxygène et de payement de dette d'oxygène qui s'observe en sortie de plongée (figure 3-62C). Dans ce cadre, la quantité d'oxygène consommée en supplément devrait, en principe, être équivalente à celle non consommée pendant la période anaérobie. Ce n'est guère le cas, la quantité d'oxygène consommée en supplément étant nettement plus petite. (figure 3-62C, figure 3-63, zones beige foncé et zones beige clair). Ce type de réponse parait général chez les animaux effectuant de longues plongées pendant lesquelles ils doivent passer à un système métabolique anaérobie. Elle a été mise en évidence non seulement chez le phoque mais également chez le canard et l'alligator. Elle pourrait être indicative d'une diminution du métabolisme durant la période anaérobie. Une diminution du métabolisme au niveau musculaire est peu probable, les animaux étant en général actifs en plongée. Elle n'aurait par contre rien de surprenant au niveau splanchnique si l'on considère l'arrêt pratiquement total de la circulation à ce niveau et le fait que l'activité métabolique d'un tissu peut dépendre dans une certaine mesure de la disponibilité de l'oxygène . Une autre possibilité serait le passage d'un système de métabolisme anaérobie de type lactate à un autre plus efficace et dans lequel le payement de la dette d'oxygène s'effectue suivant un rapport différent de celui du système lactate. Ces hypothèses attendent une étude expérimentale qui permettra de les vérifier ou de les infirmer.
Figure 3-62: Enregistrement de différents
paramètres lors d'une plongée de 12 minutes chez le phoque de Weddell.
Figure 3-63: Dette d'oxygène et payement chez un phoque gris effectuant des plongées de durée plus ou moins longue.
Les modifications de flot sanguin concomitantes à la plongée anaérobie chez le phoque sont accompagnées d'une diminution notable de la fréquence cardiaque ( figure 3-64). Un réflexe similaire a été mis en évidence chez de nombreuses autres espèces. Dans certains cas cependant cette brachycardie ne peut être démontrée. Les serpents marins par exemple maintiennent un rythme cardiaque élevé pendant la plongée; ceci pourrait être mis en rapport avec le fait qu'ils peuvent respirer par la peau et que pour profiter de l'oxygène ainsi obtenu, ils doivent maintenir leur circulation périphérique. Par ailleurs de nombreuses espèces, reptiles, oiseaux et mammifères peuvent effectuer de courtes plongées sans présenter de brachycardie. Il semble que dans ces cas, la plongée qui n'est que de courte durée, reste aérobie.
Les animaux pratiquant des plongées profondes de relativement longue durée doivent éviter les accidents de décompression que l'on peut mettre en évidence chez l'homme lors de remontées trop rapides. Ces problèmes d'aéroembolisme sont dus au fait que lors de la descente, la pression sur la cage thoracique augmente, augmentant la pression des gaz dans le poumon et dès lors la pression des gaz dissous dans le sang (O2, CO2, mais aussi et surtout N2). Si la remontée est trop rapide en fin de plongée, le N2 est brutalement libéré dans le sang à l'état gazeux. Il y a formation de bulles qui peuvent engendrer une embolie gazeuse. Comme nous l'avons vu, le phoque, à l'inverse de l'homme, possède une cage thoracique de petite dimension. Il peut par ailleurs expirer avant une plongée de façon à vider ces poumons des gaz qui s'y trouvent. Lors de la plongée, il laissera de toute façon les gaz restants dans les poumons s'échapper librement lors de la compression de la cage thoracique. Une cage thoracique de faible volume lui permet d'éviter les problèmes mécaniques qui pourraient résulter de la compression à grande profondeur. L'homme par contre va inspirer avant la plongée de façon à partir avec la réserve d'air la plus élevée possible. Il va également lutter contre la pression lors de la descente pour garder un maximum de cet air. Il risque de ce fait des problèmes d'aéroembolisme s'il ne respecte pas lors de la remontée les paliers de décompression qui lui sont prescrits. Ces paliers augmentent considérablement le temps de remontée mais permettent l'équilibration des pressions sanguines et pulmonaires sans formation de bulles.
L'homme bien que mal adapté à la plongée la pratique largement. Les techniques utilisées sont variées, allant de la plongée libre en apnée ou avec tuba jusqu'aux plongées avec équipements spéciaux tels que scaphandres alimentés depuis la surface ou encore totalement autonomes.
Le problème essentiel de la plongée en scaphandre est l'aéroembolisme (voir 4.2 ci avant).
La plongée avec tuba est soumise à deux contraintes majeures au plan respiratoire. Tout d'abord, comme nous l'avons signalé précédemment, ce tube augmente, plus ou moins largement selon sa longueur, l'espace mort anatomique du système respiratoire; ce qui, à la limite, peut empêcher toute respiration. Par ailleurs, les mouvements respiratoires deviennent rapidement difficiles par suite de l'augmentation de la pression hydrostatique. L'inspiration devra en fait se faire contre une certaine pression qui risque d'en diminuer l'ampleur (figure 3-65). Lors d'une inspiration, la dépression maximum pouvant être produite est d'environ 11 KPa, soit 112 cm H2O. La respiration d'air à pression atmosphérique devient donc impossible à partir d'environ 1,10 m de profondeur; limite à partir de laquelle le thorax reste en position d'expiration et le sujet entre en anoxie hypoxémique.
 |
|
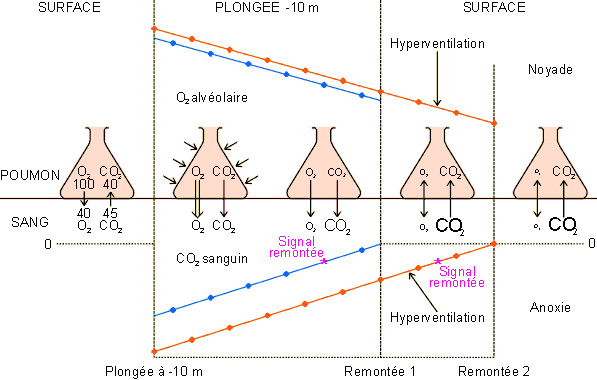
La plongée en apnée est, elle, sujette à un autre problème lié à l'effet de la pression sur les échanges alvéolaires (figure 3-66).
Chez le plongeur en apnée, le signal "remontée" est donné par une sensation de dyspnée qu'engendre l'élévation de la teneur du sang en CO2 par l'intermédiaire de chémorécepteurs particuliers. Pour prolonger sa plongée, le sujet peut passer outre les premiers signaux de dyspnée. Cette pratique peut avoir des conséquences dramatiques surtout si comme c'est souvent le cas, le plongeur hyperventile ses poumons avant le départ pour augmenter sa réserve d'oxygène. Cette pratique va aussi diminuer le CO2 alvéolaire et sanguin; ce qui, comme nous allons le voir, va augmenter le risque d'anoxie hypoxémique et peut mener le sujet à la mort. En conditions normales, les gradients de pression partielle O2 et CO2 entre le sang et l'air alvéolaire favorisent le passage de l'O2 des poumons vers le sang et le passage du CO2 du sang vers les poumons ( tableau 3-5, figure 3-66). En plongée, la pression augmente dans les poumons ce qui facilite la diffusion de l'oxygène mais freine celle du CO2. En fait, à 10 m de profondeur dans l'exemple choisi, la pression intra-pulmonaire est telle que le mouvement du CO2 est inversé: il va des poumons vers le sang plutôt que du sang vers les poumons. Le stock d'O2 au niveau des poumons diminue donc plus rapidement en profondeur qu'en surface alors que la PCO2 sanguine augmente,elle , plus rapidement. Le signal "remontée" sera donc donné trop tôt par rapport à l'O2 effectivement consommé: ce qui peut donner au plongeur inexpérimenté connaissant mal ses limites l'impression qu'il peut encore rester au fond quelque temps. A la remontée, la pression des gaz diminue rapidement dans les poumons et dans le sang. L'échange d'oxygène cesse alors, amenant l'hypoxie et un risque d'anoxie et de noyade si le plongeur n'atteint pas la surface suffisamment vite pour reprendre de l'oxygène. Ce risque est d'autant plus grand que le temps passé en plongée est important et donc que le stock d'O2 pulmonaire est bas. Il est par ailleurs largement augmenté par une hyperventilation préalable excessive. En effet, cette pratique, si elle augmente un peu la PO2 au niveau alvéolaire, diminue par contre nettement la PCO2, retardant ainsi le signal "remontée" qui risque alors d'arriver trop tard.
Figure 3-66: Les échanges respiratoires lors d'une plongée en apnée à 10 m: voir explications dans le texte.
Un autre problème de la plongée en apnée est le "blood shift" qui a rapport à l'appel de sang créé dans la cavité pleurale par une descente profonde rapide. Lors de la descente, l'air est en effet comprimé dans les poumons et ceux-ci diminuent largement de volume. La cage thoracique, plus rigide que le tissu pulmonaire, ne peut suivre le mouvement ; il se crée donc une dépression au niveau de la cavité pleurale qui, si elle est suffisamment importante et si la descente est suffisamment rapide, peut générer une stase sanguine importante avec un défaut de retour veineux pouvant entraîner la syncope et la noyade (voir aussi chapitre 2, 4.3.3).