2.2.1. La pituitaire ou complexe neuro-adťnohypophysaire
a. morphologie, ťvolution, prťsentation gťnťrale
b. hormones de l'adťnohypophyse
c. hormones de la neurohypophyse
d. contrŰle des productions du complexe neuro-adťnohypophysaire2.2.2. La thyroÔde
a. production et rŰle des hormones
b. synthŤse des hormones thyroÔdiennes
c. contrŰle de production des iodothyronines2.2.3. Les parathyroÔdes
2.2.4. őlots endocrines du pancrťas
2.2.5. Les surrťnales
2.2.6. Les gonades
2.2.7. Autres glandesa. la glande pinťale et le complexe ťpiphysaire
b. le thymus
c. le tube digestif
d. le cúur et le facteur atrial natriurťtique
2.3.1. Rťgulation hormonale de la calcťmie sanguine
2.3.2. Rťgulation hormonale de la volťmie et de la natriťmiec. le systŤme rťnine - angiotensine
d. le facteur natriurťtique auriculaire
e. autres hormones2.3.3. Rťgulation hormonale de la glycťmie
a. insuline et glucagon
b. contrŰle de sťcrťtion
c. effets de l'insuline et du glucagon
d. dťficience en insuline2.3.4. Rťactions hormonales aux stress
2.3.5. Rťgulation hormonale de la croissancea. sťcrťtion et effets de la GH
b. effets synergiques d'autres hormones
c. mues et mťtamorphoses2.3.6. ContrŰle hormonal de la gamťtogenŤse
1) Phase prťovulatoire
2) Ovulation
3) Phase postovulatoire
4) Effets des hormones produitesb. gamťtogenŤse testiculaire
c. aspects comparťs
2.4.1. Croissance, mues et mťtamorphoses
2.4.2. Iono et osmorťgulation
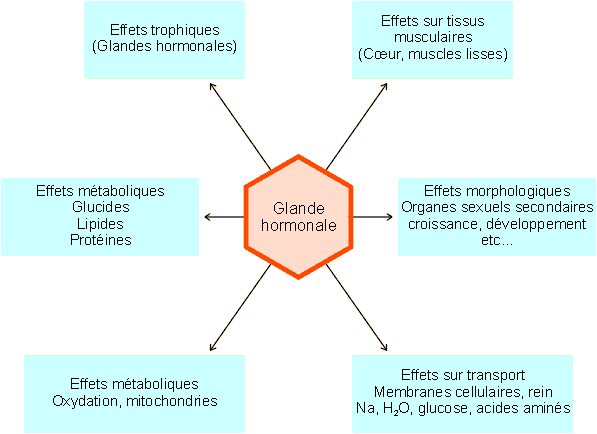
Comme nous l'avons dťjŗ signalť, les hormones sont des sťmiomolťcules sťcrťtťes par des groupes de cellules caractťristiques. Dťversťes dans le sang, elles atteignent leurs cellules - cibles aprŤs un trajet plus ou moins long dans le systŤme circulatoire. Les hormones agissent gťnťralement ŗ trŤs basse concentration (10-10 - 10-7 M) et produisent des effets extrÍmement spťcifiques soit sur tous les types cellulaires soit sur certains tissus ou organes. On distinguera ainsi des hormones ŗ action ubiquitaire et des hormones ŗ action restreinte. Les hormones agissant sur tous ou sur de nombreux types cellulaires ont en gťnťral des effets mťtaboliques : rťgulation du mťtabolisme du glucose, des lipides et/ou des protťines, rťgulation du mťtabolisme oxydatif (par exemple : hormones de croissance, thyroÔdienne, insuline, etc.Ö). Les autres hormones n'agissent en gťnťral que sur un ou quelques tissus particuliers ; elles peuvent avoir des effets trophiques (induisant la sťcrťtion d'autres hormones), des effets morphologiques, des effets sur le tonus des muscles lisses ou cardiaque ou sur des mťcanismes de transport (figure 9-2).
Figure 9-2 : Types d'effets hormonaux.
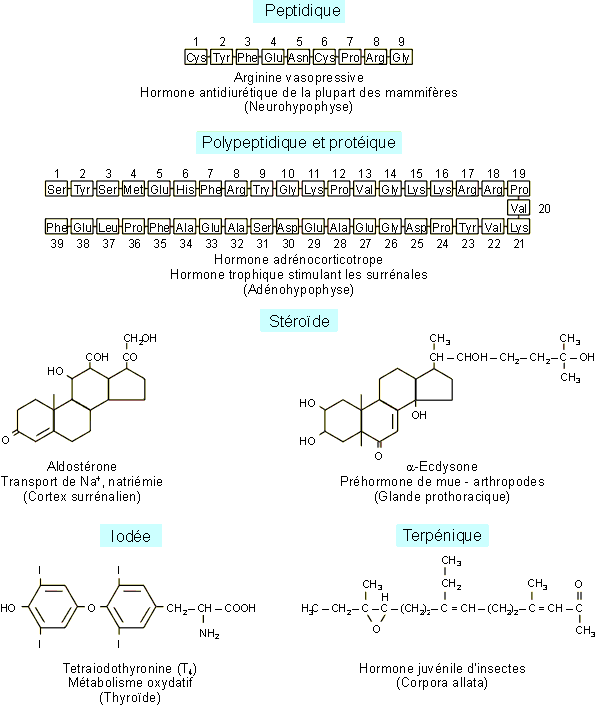
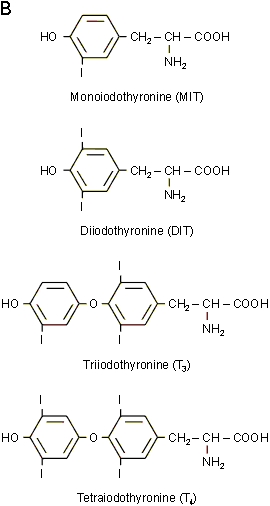
La plupart des hormones sont de nature polypeptidiques ou protťiques. Certaines d'entre elles, comme les hormones sexuelles, les hormones du cortex surrťnalien des vertťbrťs ou les hormones de mue des arthropodes, sont des stťroÔdes dťrivťs du noyau squalŤne. Quelques hormones enfin ne font pas partie de ces deux grandes catťgories. Ainsi, les hormones ŗ effets mťtaboliques de la thyroÔde des vertťbrťs sont des dťrivťs iodťs de la tyrosine et de la tyronine alors que l'hormone juvťnile des insectes est, elle, de type terpťnique (figure 9-3).
Figure 9-3 : Nature chimique des hormones ŗ travers quelques exemples.
Les hormones paraissent toutes agir par l'intermťdiaire de l'association avec un rťcepteur spťcifique de la cellule - cible. Les rťcepteurs sont soit membranaires pour les hormones ne pouvant passer la membrane plasmique (hormones peptidiques, protťiques, adrťnaline de la mťdullaire surrťnalienne) soit cytoplasmiques pour les hormones liposolubles pouvant accťder aisťment par diffusion au milieu intracellulaire (hormones stťroÔdes et thyroÔdiennes iodťes). Les problŤmes relatifs ŗ la rťception et ŗ la signalisation intracellulaire des messages hormonaux seront envisagťs en dťtail plus loin (cfr. sections 4 et 5).
Les hormones ont ťtť beaucoup plus ťtudiťes chez les vertťbrťs que chez les invertťbrťs. Il n'existe en fait pour ces derniers que quelques ťtudes pouvant ŗ l'heure actuelle Ítre intťgrťes dans des ensembles cohťrents concernant essentiellement la croissance et la mue. Nous nous concentrerons donc essentiellement sur les glandes endocrines et sur quelques exemples de rťgulations hormonales chez les vertťbrťs.
Les vertťbrťs possŤdent une dizaine de structures pouvant Ítre considťrťes comme glandes endocrines. Certaines d'entre elles sont homologues chez les diffťrentes espŤces et produisent des hormones souvent de structures chimiques identiques ou trŤs proches et ayant des effets similaires sur les mÍmes systŤmes - cibles (figure 9-4). L'homologie est cependant loin d'Ítre de rŤgle. Ainsi les parathyroÔdes n'existent pas chez les poissons et il n'y a pas de cellules a dans les Ólots endocrines pancrťatiques des urodŤles. De mÍme, certains tissus qui pourraient avoir des activitťs de type hormonales chez les poissons comme les corpuscules stannieux ou l'urophyse sont absents dans les autres groupes.
Figure 9-4 : Evolution du systŤme hormonal chez les vertťbrťs. Il existe de trŤs nombreuses homologies entre le systŤme endocrinien de l'homme et d'un poisson tťlťostťen. Il existerait chez les poissons un systŤme supplťmentaire, celui de l'urophyse, quoique son rŰle comme glande hormonale reste peu clair. Dťtails dans le texte.
La prťsentation des glandes hormonales sera dŤs lors axťe sur le systŤme endocrinien des mammifŤres en ne considťrant pour les diffťrentes glandes que les aspects comparťs les plus marquants. Nous renvoyons le lecteur intťressť par une ťtude comparative plus dťtaillťe ŗ l'excellent ouvrage de Norris "Vertebrate endocrinology".
2.2.1. La pituitaire ou complexe
neuro-adťnohypophysaire
a. morphologie, ťvolution, prťsentation gťnťrale
La glande pituitaire est prťsente dans le cerveau de tous les vertťbrťs, des
poissons aux mammifŤres. Elle peut Ítre considťrťe comme l'une des glandes
endocrines les plus importantes par le nombre d'hormones qu'elle secrŤte et
dŤs lors par la variťtť des activitťs biologiques qu'elle contrŰle.
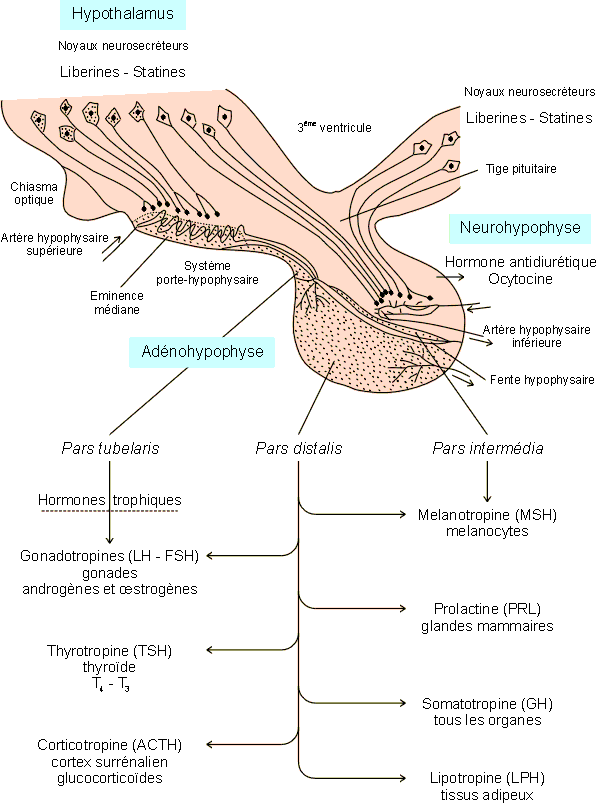
Elle est constituťe de deux parties morphologiquement et embryologiquement distinctes. La neurohypophyse, d'une part, qui, comme son nom l'indique, est d'origine nerveuse et dťrive embryologiquement du plancher du diencťphale (thalamus, hypothalamus). L'adťnohypophyse, d'autre part, qui est dťrivťe d'une ťvagination de la voŻte palatine et qui vient se fixer, lors du dťveloppement embryonnaire, ŗ l'avant de la neurohypophyse. Dans ce cadre, l'adťnohypophyse est parfois appelťe lobe antťrieur de l'hypophyse, la neurohypophyse devenant le lobe postťrieur ou posthypophyse. L'hypophyse reste reliťe ŗ l'hypothalamus dans le cerveau par la tige pituitaire, l'ensemble ťtant souvent considťrť comme un tout fonctionnel : le complexe hypothalamo-hypophysaire (figure 9-5). La neurohypophyse reÁoit ainsi les prolongements des cellules neurosecrťtrices de diffťrents noyaux hypothalamiques. L'adťnohypophyse par contre se prolonge vers l'hypothalamus en un systŤme porte-hypophysaire. Celui-ci assure le transport des productions hypothalamiques et donc le contrŰle par le cerveau des productions hormonales du lobe antťrieur( voir d ci-aprŤs).
La paroi postťrieure de l'ťbauche buccale, accolťe ŗ la neurohypophyse, subit un dťveloppement souvent trŤs modťrť. Elle va constituer un lobe intermťdiaire entre la partie avant de l'adťnohypophyse et la neurohypophyse. On distinguera ainsi dans l'adťnohypophyse une pars intermedia d'une autre, plus importante qui la jouxte, la pars distalis. Dans ce contexte, la neurohypophyse deviendra la pars nervosa de l'hypophyse.
La pars intermedia est bien dťveloppťe chez les poissons, les amphibiens et les reptiles ; elle est absente chez les oiseaux et trŤs rudimentaire chez l'homme oý elle disparaÓt pratiquement ŗ la pubertť par incorporation de ses cellules dans le lobe antťrieur. La lumiŤre de l'invagination buccale embryonnaire (poche de Rathke) subsiste entre les pars distalis et intermedia sous la forme d'une cavitť ťtroite, la fente hypophysaire, bien visible chez les rongeurs, pratiquement inexistante chez l'homme (figure 9-5). L'adťnohypophyse comporte encore une partie morphologiquement distincte des deux autres, la pars tubelaris, se localisant vers le haut en dessous de l'ťminence mťdiane.
Figure 9-5 : Le complexe hypothalamo-hypophysaire - organisation et productions hormonales . L'adťnohypophyse (lobe antťrieur) est reliťe ŗ l'hypothalamus par le systŤme porte-hypophysaire (circulation sanguine) alors que la neurohypophyse reÁoit les terminaisons des cellules neurosecrťtrices de diffťrents noyaux hypothalamiques. Dťtails dans le texte.
b. hormones de l'adťnohypophyse
L'adťnohypophyse prťsente, dans ces diffťrentes parties, diffťrents types
cellulaires ayant fait l'objet de nombreuses classifications histologiques
souvent complexes et parfois contradictoires. Des techniques d'immunovisualisation
rťcentes permettent de classer ces cellules en diffťrents types fonctionnels,
suivant le type d'hormones qu'elles produisent, toutes ces hormones ťtant de
nature protťique.
La pars distalis et la pars tubelaris contiennent essentiellement des cellules sťcrťtant des hormones ŗ effet trophique, c'est ŗ dire stimulant la production d'autres hormones.
On y trouvera ainsi :
- des cellules gonadotropes, sťcrťtant les gonadotropines LH (hormone
lutťinisante) et FSH (hormone folliculo-stimulante) qui vont activer les
productions d'hormones sexuelles au niveau des gonades ainsi que la formation
des gamŤtes (voir plus loin 2.2.6 et 2.3.6).
- des cellules thyrťotropes dont la sťcrťtion, la TSH (hormone thyrťotrope,
thyrotropine) active les productions hormonales de la thyroÔde (voir plus loin 2.2.2).
- des cellules corticotropes produisant l'ACTH (hormone adťnocorticotrope,
corticotropine) qui active la production de la partie corticale des glandes
surrťnales (voir plus loin 2.2.5).
On y trouvera encore :
- des cellules dites lactotropes sťcrťtant la prolactine (PRL), une hormone
impliquťe chez les mammifŤres dans le dťveloppement, la stimulation des
glandes mammaires et la montťe laiteuse (voir 2.3.6).
La prolactine induit ťgalement le dťveloppement des glandes du jabot des
pigeons et la desquamation de leurs couches cellulaires internes formant le
"lait" de pigeon. Elle paraÓt ťgalement induire une tendance ŗ la
couvaison chez les oiseaux ainsi qu'un comportement maternel chez les rongeurs.
Chez les poissons, elle pourrait par ailleurs jouer un rŰle dans l'osmorťgulation
puisqu'elle paraÓt impliquťe dans le contrŰle de la prolifťration des
cellules ŗ chlorure des branchies (cfr. chapitre 1).
- des cellules somatotropes sťcrťtant l'hormone de croissance (GH) qui joue un
rŰle essentiel dans le dťveloppement pondťral de tous les organes et tissus.
La pars tubelaris est essentiellement riche en cellules gonadotropes produisant LH et FSH. Ces cellules reprťsentent ainsi chez l'homme quelque 80 % des cellules de cette zone morphologique. Les autres types cellulaires citťs ci-dessus y sont nettement plus rares et les productions de TSH, ACTH, PRL et GH proviennent essentiellement de la pars distalis qui produira ťgalement, comme nous l'avons vu, les gonadotropines LH et FSH.
La pars distalis produit encore deux facteurs polypeptidiques dont le rŰle physiologique reste ŗ dťfinir mais qui pourraient avoir un effet sur la mobilisation des graisses, d'oý leur nom de lipotropines (LPH). Ces facteurs proviennent en fait du clivage, dans les cellules corticotropes, d'une glycoprotťine, la pro-opio-mťlanocortine (POMC), qui est le prťcurseur commun de l'ACTH et des LPH ainsi que des b endorphines et des hormones mťlanotropes.
La production de b endorphines est plus importante au niveau de la pars intermedia qu'au niveau de la pars distalis. La pars intermedia produira encore ŗ partir du POMC des hormones mťlanotropes (a , b , g MSH) alors que la pars distalis produira surtout l'ACTH, chacun des lobes ayant ces propres modalitťs de clivage du prťcurseur.
Les b endorphines, agoniste des rťcepteurs aux opioÔdes (cfr. chapitre 8) possŤdent un important pouvoir analgťsique. Les effets analgťsiques centraux semblent cependant devoir Ítre davantage attribuťs ŗ la libťration de b end. par des terminaisons nerveuses qu'ŗ un relarguage hypophysaire. La b end. circulante ne passe en effet que trŤs mal la barriŤre hťmato-encťphalique et, par ailleurs, la b end. sťcrťtťe par la pars intermedia est acťtylťe et tronquťe dans sa portion C terminale par rapport ŗ la b end. neuronale, ce qui la rend peu active sur les rťcepteurs aux opioÔdes.
Les hormones mťlanotropes agissent essentiellement sur les mťlanocytes en stimulant la mťlanogenŤse (synthŤse de mťlanine) ainsi que la translocation des granules contenant la mťlanine (mťlanosomes). Une sťcrťtion de MSH provoquera donc chez l'homme un assombrissement de la peau. Chez les tťlťostťens, les mťlanocytes sont sous contrŰle d'une innervation adrťnergique. Une stimulation des rťcepteurs provoque l'agrťgation des mťlanosomes et un ťclaircissement alors qu'une stimulation b adrťnergique assure leur dispersion et l'assombrissement.
c. hormones de la
neurohypophyse
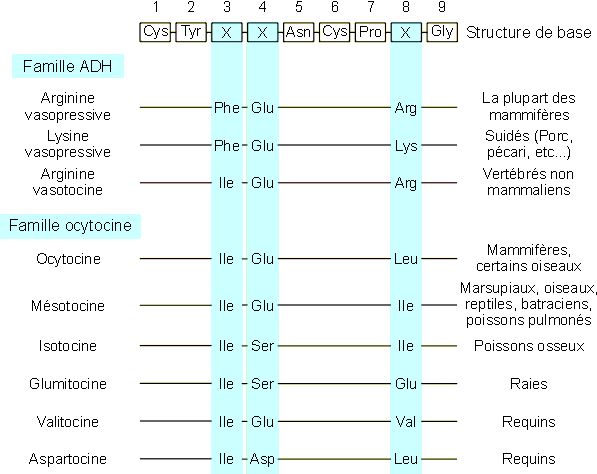
La neurohypophyse produit deux hormones prťsentant de grandes analogies
structurales : l'hormone antidiurťtique (ADH) et l'ocytocine (OT). Il s'agit en
effet dans les deux cas de peptides de 9 ou 10 acides aminťs dont la structure
gťnťrale est restťe extrÍmement conservťe chez les vertťbrťs. En ce qui
concerne l'ADH par exemple, seuls les acides aminťs 3 et 8 ont en effet ťtť
sujet ŗ modifications au cours de l'ťvolution (figure 9-6).
Figure 9-6 : Structure des hormones neurohypophysaires (ADH et OT) chez les vertťbrťs.
Les deux hormones ont une origine hypothalamique, les fibres sťcrťtrices de la neurohypophyse ťmanant presque exclusivement de neurones de deux noyaux de l'hypothalamus : le noyau paraventriculaire et, surtout, le noyau supraoptique. La destruction sťlective de ces noyaux entraÓne la dťgťnťrescence des fibres nerveuses de la tige hypophysaire et du lobe postťrieur ainsi qu'un arrÍt de la production hormonale.
L'hormone antidiurťtique est synthťtisťe sous forme d'un prťcurseur macromolťculaire (Ī 20 KDa). Elle est impliquťe essentiellement dans le contrŰle de la rťabsorption d'eau par le rein. Elle paraÓt ťgalement provoquer, ŗ doses importantes, une certaine vasoconstriction, principalement des rťgions splanchniques et rťnales. L'effet antidiurťtique de l'hormone est prťsent, plus ou moins marquť, chez tous les tťtrapodes. Chez les poissons, curieusement, seul un effet inverse, diurťtique, a ťtť dťcrit. Il est particuliŤrement spectaculaire chez les poissons pulmonťs au niveau desquels la mesotocine, une hormone de la famille de l'ocytocine, est ťgalement active. Nous envisagerons les effets de l'ADH de faÁon plus dťtaillťe en discutant le contrŰle hormonal de la volťmie sanguine (cfr. 2.3.2).
L'ocytocine induit des contractions de la musculature du tractus gťnital femelle essentiellement lors de la parturition mais aussi lors du coÔt. La sťcrťtion d'OT lors de l'accouchement est dťclenchťe par la descente du fútus et la dilatation du col de l'utťrus. L'OT est ťgalement ŗ la base du rťflexe d'ťjection du lait dťclenchť par la tťtťe, la traite ou la traction du mamelon. Chez diffťrentes espŤces non mammaliennes comme les amphibiens, l'ocytocine a des effets similaires ŗ l'hormone antidiurťtique, augmentant rapidement la permťabilitť ŗ l'eau de la peau et de la vessie. Elle stimule ťgalement le transport actif de sodium au niveau de ces ťpithťlia.
d. contrŰle des productions
du complexe neuro-adťnohypophysaire
Les productions de l'adťnohypophyse sont contrŰlťes par de nombreux facteurs
dont l'ťtude dťtaillťe sort largement du cadre de cet ouvrage. TrŤs
briŤvement, il existe un contrŰle nerveux important qui sera adrťnergique,
GABAergique ou dopaminergique selon le cas. L'hypothalamus produit par ailleurs
des facteurs stimulant (libťrines - RF : releasing factors) ou inhibant (statines
- IF : inhibiting factors) les productions hormonales. On trouvera ainsi des
prolactostatines (PRL - IF) et des prolactolibťrines (PRL - RF) rťgulant la
sťcrťtion de prolactine, des gonadostatines (LH / FSH - IF) et des
gonadolibťrines (LH / FSH - RF) rťgulant la sťcrťtion des hormones
gonadotropes, ou encore des thyrťolibťrines (TRF) stimulant la sťcrťtion des
thyrotropines, etc.Ö De nombreux autres facteurs peuvent encore intervenir.
Ainsi par exemple, le peptide intestinal vasoactif et d'autres peptides du mÍme
groupe (VIP) exerce un puissant effet stimulant sur les cellules ŗ prolactine,
notamment chez les primates et les rongeurs. Il en va de mÍme pour l'ocytocine,
l'angiotensine II et la thyrťolibťrine. Les hormones thyroÔdiennes (T3
notamment) exercent par contre un effet inhibiteur. Certains de ces effets
seront discutťs dans le cadre de l'ťtude de quelques contrŰles hormonaux (cfr.
2.3).
En ce qui concerne la neurohypophyse, le contrŰle de la production d'ocytocine est relativement mal connu, la nature des rťflexes d'ťjection du lait ou de stimulation des contractions en cours de travail restant peu claires. La rťgulation de la sťcrťtion de l'ADH sera envisagťe dans le cadre de l'ťtude du contrŰle hormonal de la volťmie (section 2.3.2).
2.2.2. La thyroÔde
a. production et rŰle des hormones
La thyroÔde des mammifŤres contient deux types de cellules endocrines
produisant chacune des hormones diffťrentes : les cellules folliculaires qui
forment les follicules thyroÔdiens synthťtisant et sťcrťtant les hormones
thyroÔdiennes iodťes. Les cellules parafolliculaires ou cellules C qui,
isolťes ou regroupťes en amas, sont localisťes entre les follicules et
sťcrŤtent essentiellement la calcitonine mais aussi deux peptides directement
apparentťs, la katacalcine et le CGRP (calcitonine - gene - regulated -
peptide) et un peu de somatostatine.
La calcitonine intervient essentiellement dans la rťgulation du mťtabolisme calcique. Son rŰle hypocalcťmiant sera envisagť de faÁon plus dťtaillťe dans le cadre de l'ťtude de quelques exemple d'effets hormonaux (cfr. 2.3.1 ci-aprŤs). Les cellules C thyroÔdiennes n'existent que chez les mammifŤres. La calcitonine est produite par les corps ultimobranchiaux depuis les poissons jusqu'aux oiseaux. Chez les tťlťostťens, un facteur hypocalcique paraÓt ťgalement prťsent dans les corpuscules stannieux (cfr. 2.3.1).
Les hormones thyroÔdiennes iodťes interviennent essentiellement dans le contrŰle du mťtabolisme oxydatif gťnťral. Elles favorisent ainsi la croissance gťnťrale et le dťveloppement. Elles sont par ailleurs indispensables ŗ la mťtamorphose de diffťrentes espŤces (tÍtard de grenouille, plie). Elles paraissent ťgalement impliquťes, ŗ cŰtť des glucocorticoÔdes, dans la smoltification du saumon. A cŰtť de ces effets sur le dťveloppement que l'on retrouve chez tous les vertťbrťs, les hormones thyroÔdienne exercent des effets mťtaboliques en relation avec la thermogenŤse spťcifiquement chez les espŤces homťothermes, oiseaux et mammifŤres (voir chapitre 12).
Aux doses physiologiques, ces hormones stimulent la production des enzymes respiratoires. Les mitochondries se dťveloppent et multiplient leurs crÍtes. Il y a augmentation de la consommation d'oxygŤne et de la production d'ATP. En l'absence d'hormones, la consommation d'oxygŤne est rťduite de moitiť et le mťtabolisme oxydatif est bas. L'hypothyroÔdisme peut ainsi conduire au nanisme, ŗ l'infantilisme ou ŗ l'idiotie (crťtinisme). L'hyperthyroÔdisme au contraire peut provoquer un gonflement des mitochondries avec augmentation importante de la consommation d'oxygŤne et du catabolisme cellulaire en mÍme temps qu'un dťcouplage des phosphorylations oxydatives. Il s'ensuit un arrÍt de croissance et de dynamisme pouvant conduire ŗ la cachexie.
b. synthŤse des hormones
thyroÔdiennes
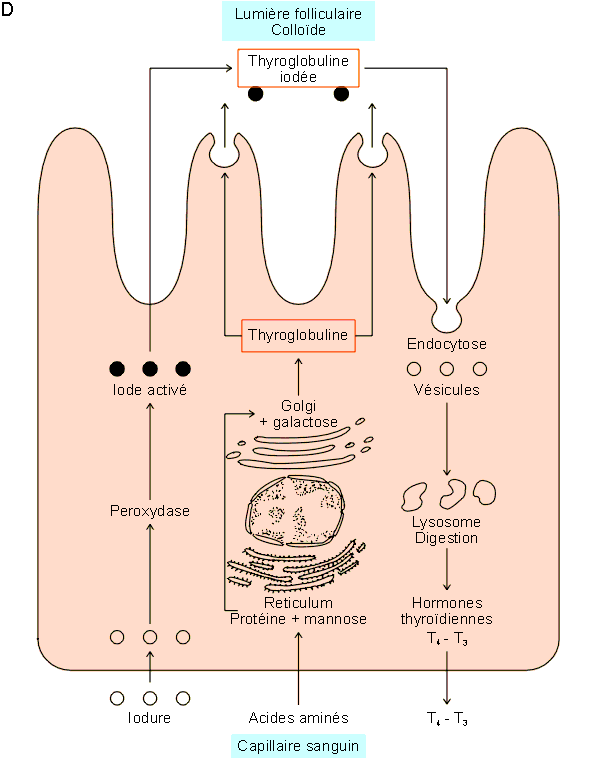
Les hormones thyroÔdiennes sont formťes ŗ partir d'une grosse protťine, la
thyroglobuline, qui fixe l'essentiel de l'iode se trouvant dans l'organisme
(chez l'homme : 8 des 9 mg d'iode total). La thyroglobuline est une
glycoprotťine synthťtisťe dans les cellules folliculaires de maniŤre
relativement complexe impliquant l'addition de parties glucidiques dans le
reticulum et le Golgi (cfr. figure 9-7D). Une fois formťe,
elle est exocytťe au pŰle apical des cellules dans le colloÔde central du
follicule oý elle est iodťe ŗ partir d'iode actif rťsultant de l'oxydation
par une peroxydase, dans les cellules folliculaires, d'iodure pompť dans le
sang. La thyroglobuline iodťe est alors endocytťe par les cellules
folliculaires et clivťe dans les lysosomes en donnant naissance aux
diffťrentes hormones thyroÔdiennes (iodothyrosines et iodothyronines) parmi
lesquels la T4 (Thyroxine) est la plus importante (figure
9-7B). Les hormones ainsi formťes peuvent alors Ítre dťversťes dans le
sang.
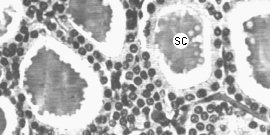
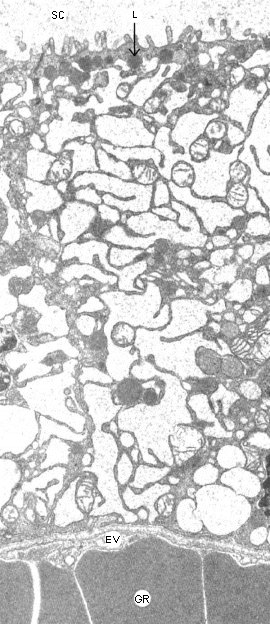
Figure 9-7ABC : Les hormones thyroÔdiennes. A : Vue en microscopie ťlectronique d'une section de glande montrant les structures folliculaires avec leur lumiŤre de substance colloÔdale (SC) et les cellules les entourant. B : Structure des hormones thyroÔdiennes. C : Vue en microscopie ťlectronique d'une cellule folliculaire. La thyro-globuline est endocytťe ŗ la face apicale et transformťe dans les lysosomes (L) en hormones thyroÔdiennes qui sont relarguťes dans le sang (EV : endothťlium vasculaire, GR : globules rouges).
Figure 9-7D : Les hormones thyroÔdiennes. Schťma explicatif des mťcanismes de synthŤse. Dťtails dans le texte.
c. contrŰle de production des
iodothyronines
La thyroÔde reÁoit une importante innervation adrťnergique, cholinergique et
peptidergique qui paraÓt pouvoir moduler l'effet de l'hormone thyrťotrope. La
nature exacte de cette modulation ainsi que son rŰle physiologique sont
cependant loin d'Ítre ťlucidťs. Le taux de T3 et T4
circulantes exerce par ailleurs un contrŰle en rťtroaction sur la production
de TSH. Les variations de tempťrature saisonniŤres paraissent ťgalement
influencer sa production, ce qui aboutit ŗ une augmentation du mťtabolisme de
base en pťriode froide et ŗ une diminution en pťriode chaude (figure
9-8).
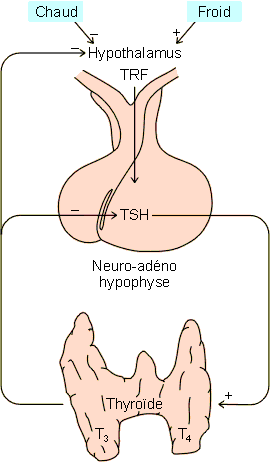 |
Figure 9-8 : ContrŰle de la production des hormones thyroÔdiennes. TRF : thyrolibťrine, TSH : hormone thyrťotrope. |
2.2.3. Les parathyroÔdes
Habituellement au nombre de 3 ou 4 chez l'homme, ces petites glandes, ne
mesurant que quelques millimŤtres, sont incrustťes sur la face postťrieure de
la thyroÔde. Elles produisent la parathormone (PTH) qui, avec la calcitonine,
joue un rŰle important dans l'ťquilibre calcique et la fixation du calcium au
niveau des os (cfr. 2.3.1). La PTH joue ťgalement un
rŰle important dans la rťgulation de la teneur en phosphate du sang en
stimulant le transport de PO4 au niveau rťnal.
Le stimulus majeur contrŰlant la sťcrťtion de PTH paraÓt Ítre le calcium. Chez le veau nouveau-nť par exemple, la quantitť de PTH sťcrťtťe est directement liťe ŗ la quantitť de calcium prťsente dans le sang, l'hypocalcťmie augmentant la sťcrťtion et l'hypercalcťmie la diminuant.
Les parathyroÔdes sont prťsentes ŗ partir des amphibiens et paraissent absentes chez les poissons. Il existe des facteurs de type PTH chez les poissons osseux ; ils seraient ťlaborťs au niveau de la pars distalis de l'adťnohypophyse (cfr. 2.3.1).
2.2.4. Ilots endocrines du
pancrťas
Le pancrťas est une glande ŗ la fois endocrine et exocrine. En temps que
glande exocrine, il participe de faÁon essentielle ŗ la digestion par sa
production de diffťrentes enzymes digestives (cfr. chapitre 5, sections 2 et 4.3.1). Le pancrťas
contient par ailleurs de minuscules amas de cellules endocrines (Ólots de
Langerhans). On y trouve essentiellement des cellules de deux types, a et b , produisant
respectivement le glucagon et l'insuline. On trouve ťgalement des cellules d qui sťcrŤtent de la somatostatine dont le rŰle est
d'inhiber toute sťcrťtion d'insuline ou de glucagon. D'autres cellules encore
produisent un polypeptide pancrťatique (PP) dont le rŰle physiologique exact
reste toujours inconnu. Il semble qu'il pourrait agir sur les processus
digestifs en stimulant notamment la sťcrťtion et la motricitť gastrique.
L'insuline et le glucagon ont des effets marquťs sur le mťtabolisme glucidique. Ces hormones vont dŤs lors jouer un rŰle essentiel dans la rťgulation du taux de glucose sanguin (glycťmie), l'insuline ťtant hypoglycťmiante et le glucagon hyperglycťmiant. Ces effets seront envisagťs de faÁon plus dťtaillťe dans la section 2.3 (cfr. 2.3.3).
2.2.5. Les surrťnales
Les glandes surrťnales se prťsentent chez l'homme comme de petites structures
pyramidales localisťes au sommet des reins. Elles sont formťes par
l'association de deux portions endocrines distinctes tant par leurs origines
embryologiques que leurs fonctions. Le cortex surrťnalien, dťrivť du
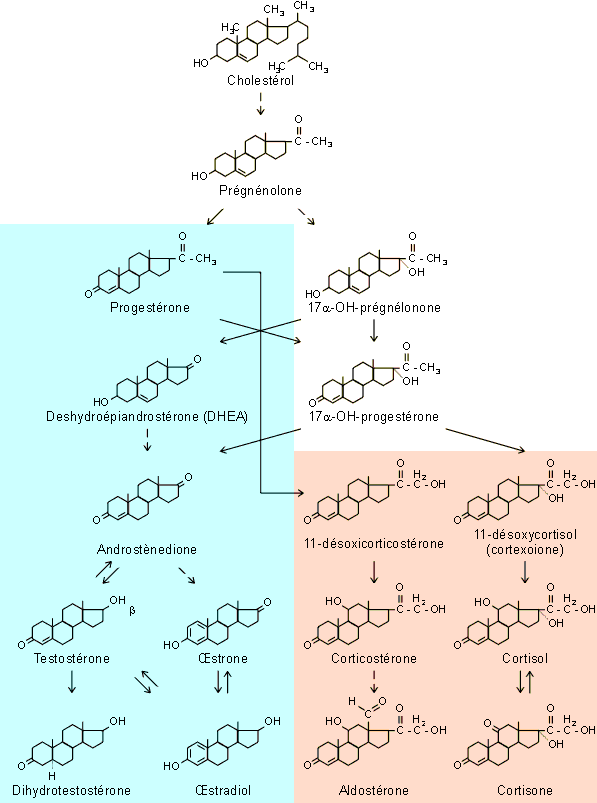
mťsoderme, produit des stťroÔdes (minťralo et glucocorticoÔdes - figure 9-9) alors que la partie mťdullaire (tissu chromaffine),
dťrivťe du neuroectoderme et plus particuliŤrement des crÍtes neurales,
ťlabore des catťcholamines. Tissus stťroÔdien et chromaffine se prťsentent
sous forme d'amas cellulaires sťparťs chez les poissons cartilagineux ;
ils se rapprochent de plus en plus au cours de l'ťvolution pour s'unir en une
seule glande chez les mammifŤres.
Les catťcholamines produites par la zone mťdullaire, essentiellement adrťnaline et noradrťnaline mais aussi dopamine, vont avoir un rŰle important dans les rťactions aux stress (cfr. section 2.3.4).
Les diffťrents stťroÔdes sont produits dans des zones distinctes de la rťgion corticale. La plus extťrieure (zone glomťrulťe) produit des minťralocorticoÔdes dont l'aldostťrone (figure 9-9) est certainement le plus puissant et le plus important (95 % de la production totale chez l'homme). L'aldostťrone agit sur la rťabsorption rťnale de sodium et joue ainsi un rŰle important dans le contrŰle de la natriťmie sanguine et de la balance hydrominťrale de l'organisme (cfr. section 2.3.2). Les zones internes (fasciculťe et rťticulťe) produisent essentiellement des glucocorticoÔdes (cortisol, cortisone et corticostťrone - figure 9-9 - seul le cortisol est sťcrťtť en quantitťs notables chez l'homme). Ces hormones exercent des effets mťtaboliques notables. Elles interviendront notamment dans la production de glucose et d'acides gras lors du stress ainsi que dans la rťgulation ŗ long terme de l'activitť du mťtabolisme glucidique, lipidique et protťique, favorisant ainsi le fonctionnement normal de l'organisme (cfr. sections 2.3.4 et 5). Ces effets mťtaboliques ne sont marquťs que chez les oiseaux et surtout les mammifŤres. Chez les espŤces non mammaliennes, cortisol et corticostťrone ont avant tout des effets sur la balance hydrominťrale par activation de la Na+/K+ ATPase au niveau des tissus transporteurs d'ions (branchies, reins, glandes ŗ sel). Ils peuvent agir dans ce cadre indťpendamment de ou en synergie avec l'aldostťrone.
La zone rťticulťe ťlabore ťgalement de petites quantitťs d'hormones sexuelles : les gonadocorticoÔdes. On trouve parmi celles-ci des androgŤnes et des ústrogŤnes. Normalement la production d'androgŤnes prťdomine. Leurs effets virilisants sont en gťnťral peu visible chez l'homme adulte oý la testostťrone testiculaire prťdomine largement. Ces effets sont toutefois non nťgligeables chez le jeune homme, avant la pubertť. Ils pourraient par ailleurs Ítre dťterminants dans le dťveloppement de la libido et d'une certaine masculinisation chez la femme (augmentation de pilositť et hypertrophie clitoridienne).
Chez le fútus et le nouveau-nť des primates, on trouve une zone particuliŤre trŤs dťveloppťe qui produit un androgŤne particulier en grandes quantitťs : la deshydroťpiandrostťrone (DHEA). Cette zone, dite fútale, cesse progressivement de fonctionner aprŤs la naissance et dťgťnŤre rapidement. Chez l'homme, une petite production de DHEA est maintenue au niveau de la zone rťticulťe jusqu'ŗ l'‚ge de 20 ans. Le rŰle physiologique de cette hormone reste obscur. Chez le fútus, elle pourrait servir de prťcurseur pour la synthŤse d'ústrogŤnes par le placenta (cfr. figure 9-9).
Figure 9-9 : Les hormones stťroÔdes. Produites par les gonades et les surrťnales, elles ont pour prťcurseur commun le cholestťrol. Le groupe des hormones sexuelles (bleu) comprend des hormones femelles (progestťrone, ústrone, ústradiol) et m‚les (testostťrone et dihydrotesto-stťrone). Elles sont toutes formťes dans les gonades ŗ l'exception de l'hormone m‚le deshydroťpi-androstťrone (DHEA) qui est synthťtisťe par les surrťnales. Les autres hormones surrťnaliennes (beige) comprennent des minťralocorticoÔdes (aldostťrone) et des glucocorticoÔdes (cortisol, cortisone, corticostťrone). Dťtails dans le texte.
2.2.6. Les gonades
Les hormones dites sexuelles produites par les gonades sont toutes de nature
stťroÔde. Elles sont ťlaborťes par les ovaires chez la femelle et les
testicules chez le m‚le.
Les ovaires synthťtisent deux grands types d'hormones "femelles" : les ústrogŤnes et la progestťrone. Les premiŤres provoquent la maturation des organes gťnitaux et l'apparition des caractŤres sexuels secondaires femelles (cfr. 2.3.6). La progestťrone est plus particuliŤrement impliquťe dans la prťparation du systŤme reproducteur ŗ la rťception d'un úuf fťcondť et ŗ sa gestation (cfr. 2.3.6). Elle a ťgalement des effets sur la production des glandes mammaires.
Les testicules sťcrŤtent en particulier la testostťrone, hormone "m‚le" qui suscite la maturation des organes gťnitaux masculins, l'apparition des caractŤres sexuels secondaires et l'ťmergence de la libido. Elle est par ailleurs nťcessaire ŗ la production de spermatozoÔdes ainsi qu'au bon fonctionnement des organes gťnitaux chez le m‚le adulte (cfr. 2.3.6).
2.2.7. Autres glandes
A cŰtť de ces glandes importantes, il convient d'en citer rapidement quelques
autres tels la glande pinťale, le thymus, le tube digestif ou le cúur.
a. la glande pinťale et le
complexe ťpiphysaire
La glande pinťale est, chez les mammifŤres et les oiseaux, une petite
structure accrochťe au toit du troisiŤme ventricule, dans le diencťphale.
Chez les poissons, les amphibiens et diffťrents reptiles (lťzards notamment),
cette structure est associťe ŗ deux autres projections du troisiŤme
ventricule, l'organe pariťtal ou parapinťal et la paraphyse. La pinťale se
retrouve chez tous les vertťbrťs ŗ l'exception des crocodiliens, les deux
autres formations ne sont guŤre bien dťveloppťes que chez quelques
groupes : l'organe parapinťal chez les cyclostomes et les lťzards, la
paraphyse chez les amphibiens. La pinťale contient des cellules sťcrťtrices
dont la seule production notable paraÓt Ítre la mťlatonine qui exerce chez de
nombreuses espŤces des effets sur le comportement sexuel et les gonades,
l'alimentation, le sommeil, la sťcrťtion des gonadotropines de l'adťnohypophyse
et peut Ítre aussi sur la production d'autres hormones trophiques.
Le rŰle physiologique de la pinťale reste obscur. Elle reÁoit des voies visuelles des influx relatifs ŗ l'intensitť et ŗ la durťe de l'ťclairement. Elle pourrait dŤs lors Ítre impliquťe, via la mťlatonine, dans les voies responsables de diffťrents processus physiologiques rythmiques, en relation avec les cycles chronobiologiques.
b. le thymus
Le thymus est une glande souvent volumineuse chez le jeune et qui s'atrophie
avec l'‚ge. Chez tous les vertťbrťs, sauf les mammifŤres, il s'agit d'un
organe pair situť en arriŤre de la tÍte, dorsalement chez les poissons et les
amphibiens, ventralement et dans le cou chez les reptiles et les oiseaux. Chez
les mammifŤres, il se prťsente comme un organe impair, formť de deux lobes
contigus coiffant le sommet du cúur, dans la cage thoracique.
Le thymus joue un rŰle important comme organe lymphoÔde (lieu de maturation et de diffťrentiation des cellules leucocytaires). Il joue ťgalement un rŰle fondamental dans le fonctionnement du systŤme immunitaire par la production d'hormones tels que thymosine, thymaline, thymopoÔťtine ou facteur thymique humoral (THF). Ces substances interviennent en effet plus particuliŤrement dans le contrŰle de la maturation des lymphocytes T et de l'activitť de diffťrentes phases de la rťponse immunitaire (cf. chapitre 10).
Les productions hormonales thymiques mettent bien en ťvidence les interrelations existant entre le systŤme immunitaire et le systŤme endocrinien. Lymphocytes, macrophages et leucocytes montrent par ailleurs des productions immunorťactives d'ACTH, d'endorphines, de TSH ou de GH. Ces hormones sont identiques ŗ ou sont trŤs proches de celles produites par les glandes endocrines et peuvent activer les mÍmes rťcepteurs. Les relations immuno - endocriniennes seront envisagťes plus en dťtail au chapitre 10 concernant les mťcanismes de dťfense des organismes.
c. le tube digestif
Comme nous l'avons dťjŗ signalť, le tube digestif est le siŤge, en divers
endroits, d'une production hormonale importante, essentiellement liťe ŗ la
rťgulation de la fonction digestive. Le rŰle de ces hormones a ťtť abordť
au chapitre 5 dans le cadre de l'ťtude de cette
fonction ; nous n'y reviendrons donc pas ici.
d. le cúur et le facteur
atrial natriurťtique
La dťmonstration de la sťcrťtion d'un facteur hormonal par le cúur et plus
particuliŤrement par le tissu auriculaire est relativement rťcente puisqu'elle
date des annťes 1970. On a en effet pu montrť ŗ l'ťpoque qu'un extrait de
tissu auriculaire injectť dans la circulation exerÁait un puissant effet
diurťtique et natriurťtique au niveau rťnal et manifestait un pouvoir
relaxant, hypotenseur, au niveau de la circulation gťnťrale. Le facteur
hormonal en cause, produit par certains myocytes du tissu auriculaire, fut
isolť pour la premiŤre fois en 1985 et fut appelť facteur atrial
natriurťtique (ANF). Il a maintenant ťtť mis en ťvidence chez tous les
vertťbrťs, des poissons aux mammifŤres. Il s'agit d'un peptide de 27 ou 28
acides aminťs de structure assez bien conservťe (27 chez l'anguille, 28 chez
le rat et l'homme ; la diffťrence entre le peptide de l'homme et du rat ne
porte que sur l'acide aminť en position 12).
Le facteur dťclenchant la production de l'ANF paraÓt Ítre la distension des oreillettes. L'hormone produite va induire une vasodilatation, une augmentation de la diurŤse, une rťduction de sťcrťtion d'aldostťrone et une suppression de la sťcrťtion de rťnine (cfr. section 2.3.2).
2.3.1. Rťgulation hormonale de
la calcťmie sanguine
Le calcium est un ťlťment majeur de la structuration des os et des dents. Il
intervient par ailleurs dans toute une sťrie de processus tels que contraction
musculaire, exocytose ou signalisation intracellulaire. Sa concentration
sanguine (Ī 2,5 mM chez les mammifŤres) doit donc
Ítre rťgulťe avec prťcision. Líhomťostasie du calcium sanguin rťsulte
essentiellement des mouvements de l'ion au niveau osseux, urinaire et intestinal
(figure 9-10). Trois substances vont intervenir dans le
contrŰle de ces mouvements : la parathormone, la calcitonine et la vitamine D3.
Figure 9-10 : Les ťlťments de l'homťostasie du calcium sanguin chez l'homme. CT : calcitonine Ė PTH : parathormone Ė Vit D3, 25 OH D3 et 1-25 (OH)2 D3 : vitamine D3 ou cholťcalcifťrol et dťrivťs hydroxylťs. Dťtails dans le texte.
La vitamine D3 ou cholťcalcifťrol peut Ítre obtenue par apport alimentaire ou synthťtisťe par irradiation UV du 7-deshydrocholestťrol. Les amphibiens, les reptiles, les oiseaux et les mammifŤres possŤdent ainsi des zones rťceptrices aux UV au niveau desquels s'effectue la synthŤse (peau chez l'homme, pattes chez les oiseaux, oreilles chez les lapins). Les poissons paraissent capables d'ťlaborer la vitamine sans l'aide des UV. Une fois formťe ou assimilťe, la Vit D3 est transportťe par une protťine sťrique au niveau du foie oý elle peut Ítre hydroxylťe en 25-hydroxycholťcalcifťrol. Elle sera alors transportťe du foie au rein pour y subir une nouvelle hydroxylation soit en C-1 pour former un produit actif, le 1-25 dihydroxycholťcalcifťrol (1-25 (OH)2 D3), soit en C-24 pour former un produit inactif. Les effets combinťs de diffťrents modulateurs sur les enzymes impliquťes, la 1-hydroxylase et la 24-hydroxylase, vont permettre de contrŰler la concentration du mťtabolite actif. Ainsi par exemple, une calcťmie basse augmente la production de 1-25 (OH)2 D3. Cet effet implique une activation de la 1-hydroxylase et une inhibition de la 24-hydroxylase ; une calcťmie ťlevťe a l'effet inverse. Ces effets paraissent dus aux modifications de production de parathormone qu'entraÓnent les variations de la calcťmie ; la parathormone stimule en effet directement la synthŤse de la 1-hydroxylase. Une phosphatťmie basse augmente ťgalement la production de 1-25 (OH)2 D3. Contrairement ŗ ce que l'on observe pour l'hypocalcťmie, cet effet ne parait pas faire intervenir la PTH.
Le 1-25 (OH)2 D3 peut par ailleurs exercer une inhibition en rťtroaction de sa propre synthŤse par rťpression de la 1-hydroxylase et induction de la 24-hydroxylase. Il est d'autre part intťressant de remarquer que les hormones femelles (ústrogŤnes), via l'augmentation de prolactine qu'elles entraÓnent, augmentent l'activitť 1-hydroxylasique. Ce processus est particuliŤrement marquť chez les oiseaux oý il intervient au moment de la ponte, dans la calcification de la coquille de líúuf.
Le mťtabolite actif de la vitamine D3 joue un rŰle essentiel dans le contrŰle de l'absorption intestinale de calcium qu'il active largement. Le mťcanisme impliquť dans cette activation reste ŗ l'heure actuelle obscur. Le 1-25 (OH)2 D3 paraÓt stimuler ŗ la fois l'influx calcique au pŰle apical des entťrocytes et l'efflux au pŰle basal. Il induit ťgalement la formation dans les cellules intestinales d'une protťine fixant le calcium, la calbindine D, qui pourrait intervenir dans le transport intracellulaire de l'ion du pŰle muqueux au pŰle sťreux.
La parathormone, produite par les glandes parathyroÔdes (cfr. 2.2.3 ci-avant), entraÓne une mobilisation du calcium osseux 1) par activation de la rťsorption osseuse par les ostťoclastes ; 2) par augmentation de la rťabsorption de l'ion au niveau rťnal. La PTH apparaÓt donc dans ce cadre comme une hormone hypercalcťmiante.
La calcitonine, produite par les cellules parafolliculaires (cellules C) de la thyroÔde, a par contre un effet hypocalcťmiant. L'hormone agit essentiellement en inhibant l'activitť de rťsorption osseuse des ostťoclastes. Alors que le mťcanisme d'activation des ostťoclastes par la PTH paraÓt indirect et reste peu clair, le mťcanisme d'inhibition de la rťsorption par la CT est relativement bien connu. L'hormone agit en inhibant la formation et la libťration des enzymes lysosomiaux des ostťoclastes qui normalement lysent la trame osseuse. La CT inhibe par ailleurs lťgŤrement la rťabsorption de calcium au niveau rťnal chez certaines espŤces, ce qui contribue ŗ son effet hypocalcťmiant. L'importance physiologique des effets rťnaux de la CT chez l'homme reste trŤs discutťe ŗ l'heure actuelle.
Les glandes parathyroÔdes paraissent absentes chez les poissons qui, de plus, sont assez insensibles ŗ la parathormone ou ŗ des extraits de parathyroÔdes de tťtrapodes. Chez les tťlťostťens, des facteurs hypercalcťmiants semblent prťsents dans l'adťnohypophyse. Un de ceux-ci pourrait Ítre la prolactine. Un facteur hypocalcťmiant de type calcitonine paraÓt prťsent dans les corps ultimobranchiaux. Un autre agent hypocalcique (hypocalcine ou stanniocalcine) a par ailleurs ťtť mis en ťvidence dans les corpuscules stannieux. Chez les tťtrapodes jusqu'aux oiseaux, les glandes parathyroÔdes sont prťsentes et produisent une hormone hypercalcťmiante de type parathormone. Elles restent plus ou moins associťes aux corps ultimobranchiaux qui eux produisent une hormone hypocalcťmiante de type calcitonine.
2.3.2. Rťgulation hormonale de
la volťmie et de la natriťmie
Le maintien d'un volume liquidien et d'une osmolaritť constants est d'une
importance capitale pour le bon dťroulement des activitťs cellulaires (cfr. chapitre 1). De nombreuses structures dťvolues
partiellement ou exclusivement ŗ ce rŰle sont dŤs lors apparues au cours de
l'ťvolution (branchies, glandes ŗ sel, rein, intestin : cfr. chapitre 1). Diffťrents facteurs hormonaux
participent au contrŰle de l'activitť de ces structures et vont donc concourir
au maintien du volume, de la teneur en NaCl et de l'osmolaritť sanguine. Les
principaux facteurs impliquťs sont l'hormone antidiurťtique, l'aldostťrone
(chez les mammifŤres, souvent avec la corticostťrone ou le cortisol chez les
autres tťtrapodes, cfr. 2.2.5), le facteur natriurťtique
auriculaire et le systŤme rťnine - angiotensine.
a. l'hormone antidiurťtique
1) Effets biologiques
Nonapeptide sťcrťtť au niveau de la neurohypophyse (figure
9-6, 2.2.1.c), l'ADH a essentiellement pour effet
d'augmenter la permťabilitť ŗ l'eau de la paroi du tube collecteur du rein (figure 9-11A). Il en rťsulte une augmentation de la
rťabsorption d'eau et dŤs lors une diminution de la diurŤse. Il semble que
l'hormone agisse en favorisant l'ouverture d'aquaporines (canaux ŗ eau ou
hydrophores) ŗ la face apicale des cellules. A doses ťlevťes, l'ADH exerce de
plus un lťger effet vasopresseur par augmentation du tonus des muscles lisses
des vaisseaux. Ce fut en fait son premier rŰle dťcouvert d'oý le nom de
vasopressine donnť ŗ l'hormone des mammifŤres (figure 9-6,
section 2.2.1.c). La vasopressine mammalienne paraÓt
par ailleurs capable de stimuler la production d'aldostťrone du cortex des
surrťnales, ce qui contribue ŗ faciliter la rťsorption rťnale de NaCl et
dŤs lors celle de l'eau (voir b, ci-aprŤs).
 |
Figure 9-11A : Hormone antidiurťtique chez les mammifŤres. Effet sur la conductivitť hydraulique de la paroi du tubule collecteur de rein de lapin. |
2) ContrŰle de production
Les facteurs contrŰlant la sťcrťtion d'ADH ont ťtť ťtudiťs
essentiellement chez les mammifŤres. Chez le rat comme chez l'homme, la
sťcrťtion est affectťe par des modifications d'osmolaritť ou de volume
sanguin (figure 9-11B). Ce dernier paramŤtre est
vraisemblablement perÁu par les mťcanorťcepteurs carotidiens et aortiques
enregistrant la pression artťrielle (cfr. chapitre
2). Les variations d'osmolaritť seraient perÁues essentiellement au niveau
d'osmorťcepteurs hypothalamiques. Pour une osmolaritť infťrieure ŗ 280 mOsm/kg,
la concentration en ADH circulante est si faible qu'elle ne peut Ítre mesurťe
de faÁon fiable ; au-delŗ de ce seuil, la production d'hormone augmente trŤs
rapidement et proportionnellement ŗ l'osmolaritť plasmatique. Ainsi, une
augmentation de 1 % seulement de l'osmolaritť sanguine suffit pour entraÓner
un accroissement de concentration en ADH de 1 pg/ml, quantitť suffisante pour
modifier significativement le dťbit urinaire. Quand l'osmolaritť plasmatique
atteint 295 mOsm/kg, la vasopressinťmie est suffisante (Ī
5 pg/ml) pour provoquer une antidiurťsie maximale (figure
9-11B). Les osmorťcepteurs paraissent sensibles ŗ la variation d'osmolaritť
per se plutŰt qu'ŗ l'augmentation de concentration de l'un ou l'autre
solutť sanguin. Une augmentation d'osmolaritť rťalisťe par injection de
mannitol ou de saccharose est en effet tout aussi efficace que le NaCl
hypertonique pour activer la sťcrťtion d'hormone.
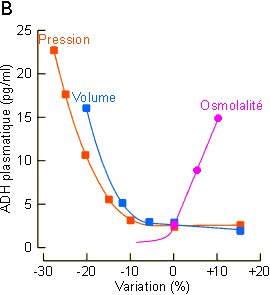 |
Figure 9-11B : Hormone antidiurťtique chez les mammifŤres. Comparaison des effets des changements d'osmolaritť, de pression et de volume sanguin sur la teneur plasmatique en ADH. |
En cas de dťshydratation, les diffťrents facteurs de contrŰle (osmolaritť, volume, pression) sont clairement liťs : une baisse de volume et donc de pression artťrielle correspond en effet ŗ une augmentation d'osmolaritť. Ils peuvent cependant agir indťpendamment. Ainsi, l'hypovolťmie rťsultant d'une hťmorragie peut induire une libťration de vasopressine mÍme si le plasma est maintenu hypoosmotique. De mÍme l'injection d'une solution hyperosmotique de NaCl ou de mannitol induit une libťration d'hormone alors qu'il n'y a pas de diminution du volume sanguin.
Curieusement, le froid et l'alcool ont des effets inhibiteurs sur la production de vasopressine, ce qui peut rendre compte de leurs effets diurťtiques. A l'inverse et tout aussi curieusement, la sollicitation du centre du vomissement de mÍme que la traction des intestins induit une production de vasopressine. Ce dernier effet peut rendre compte de l'importante rťtention d'eau qui est couramment observťe lors d'une intervention chirurgicale au niveau du tractus gastro-intestinal.
b. l'aldostťrone
1) Effets biologiques
StťroÔde produit par le cortex de la surrťnale, l'aldostťrone (figure 9-9) induit essentiellement une augmentation de la
rťabsorption de Na+ au niveau rťnal. Cet effet parait liť ŗ une
augmentation de l'activitť Na+/K+ ATPasique. On
considŤre par ailleurs que l'hormone peut intervenir directement dans le
contrŰle de la kaliťmie en stimulant la sťcrťtion rťnale du K+
lorsque sa concentration sanguine est trop ťlevťe.
L'effet de l'aldostťrone sur la Na+/K+ ATPase paraÓt assez gťnťral. Il existe chez les tťtrapodes non seulement au niveau du rein mais aussi de l'intestin, notamment dans sa portion colonique. On le trouve ťgalement au niveau de la peau et de la vessie chez les amphibiens, du cloaque chez les oiseaux, etc.Ö Il est donc essentiellement marquť au niveau des ťpithťlia transportant du sodium. Rappelons que chez les espŤces non mammaliennes, les glucocorticoÔdes, et en particulier le cortisol ont des effets similaires ; leur rŰle majeur ťtant d'intervenir au cŰtť ou ŗ la place de l'aldostťrone dans le contrŰle des mouvements de sodium (cfr. 2.2.5).
2) ContrŰle de production
Diffťrents facteurs interviennent dans le contrŰle de la production
d'aldostťrone.
La diminution de concentration en NaCl de mÍme que l'augmentation en KCl du sang induit une augmentation de sťcrťtion par activation de l'axe fonctionnel hypothalamus - hypophyse Ė surrťnales (via l'ACTH). L'ACTH paraÓt cependant agir de faÁon nettement plus importante sur la production des glucocorticoÔdes que sur celle des minťralocorticoÔdes. Les stimuli nerveux intervenant dans le contrŰle des productions du cortex surrťnalien restent ŗ l'heure actuelle peu clairs. Deux autres facteurs interviennent de faÁon importante dans le contrŰle de la production d'aldostťrone : l'angiotensine produite par le systŤme rťnine - angiotensine (SRA) et le facteur natriurťtique auriculaire (ANF).
c. le systŤme rťnine -
angiotensine
Ce systŤme apparaÓt comme le plus important intervenant dans le contrŰle de
la production d'aldostťrone. Il intervient ťgalement de faÁon majeure dans le
contrŰle de la volťmie. Il participe d'un axe hormonal impliquant le rein et
le foie.
Le fonctionnement du SRA implique tout d'abord une sťcrťtion de rťnine par un appareil juxtaglomťrulaire situť ŗ l'entrťe du glomťrule chez les mammifŤres. Cet appareil est constituť de deux parties distinctes (figure 9-12A). La premiŤre se situe sur le vaisseau affťrent et contient des cellules dites juxtaglomťrulaires qui sont les seules ŗ pouvoir sťcrťter la rťnine. Elles paraissent essentiellement sensibles ŗ la volťmie et dŤs lors ŗ la pression artťrielle qui serait enregistrťe par des barorťcepteurs se trouvant directement dans l'organe. La seconde, la Macula densa, se situe sur la partie distale du nťphron jouxtant le vaisseau affťrent. Les cellules de la Macula densa sont sensibles ŗ la concentration en sodium du tubule rťnal et peuvent induire, en fonction de ce paramŤtre, une production de rťnine des cellules juxtaglomťrulaires. Les deux stimuli efficaces, diminution de pression artťrielle (volume) et diminution de teneur en Na du tubule, peuvent agir simultanťment ou indťpendamment l'un de l'autre.
Chez les autres vertťbrťs, une Macula densa structurťe ne se trouve que chez les oiseaux. On retrouve par ailleurs au niveau du rein des cellules capables de sťcrťter la rťnine chez toutes les espŤces jusqu'aux poissons tťlťostťens, y compris ceux ayant des reins aglomťrulaires. Les cyclostomes ne paraissent pas possťder de SRA fonctionnel. La situation est moins claire chez les poissons cartilagineux oý des ťtudes rťcentes indiquent chez certaines espŤces la prťsence de structures similaires ŗ la Macula densa et aux cellules juxtaglomťrulaires. Bien que des ťtudes physiologiques restent nťcessaires, il semble que les sťlaciens au moins puissent possťder un systŤme comparable au SRA des mammifŤres.
Le SRA implique par ailleurs la production par le foie d'une glycoprotťine, l'angiotensinogŤne, qui est dťversťe dans le sang. La synthŤse de cette protťine peut Ítre stimulťe notamment par diffťrents stťroÔdes tels les ústrogŤnes et les glucocorticoÔdes.
La rťnine dťversťe dans le sang par les cellules juxtaglomťrulaire va produire par hydrolyse de l'angiotensinogŤne, un dťcapeptide, l'angiotensine I qui sera lui-mÍme converti par une enzyme dite de conversion en un octapeptide, l'angiotensine II (ou hypertensine) ou un heptapeptide, l'angiotensine III.
Figure 9-12A : Le systŤme rťnine - angiotensine. Localisation de l'appareil juxtaglomťrulaire, de la Macula densa et formation des angiotensines. CJG : cellules juxtaglomťrulaires, EC : enzyme de conversion , MD : Macula densa.
Les trois peptides formťs (angiotensine I, II et III) ont diffťrents effets biologiques dont les plus importants sont ceux de l'angiotensine II (figure 9-12BC).
L'angiotensine I peut apparemment faciliter la production de catťcholamines de la mťdullosurrťnale. Elle aurait ainsi essentiellement des effets vasoconstricteurs qui paraissent cependant rester de peu d'importance physiologique. L'angiotensine II a par contre des effets vasoconstricteurs marquťs. Elle induit par ailleurs la production d'aldostťrone de la zone glomťrulťe du cortex surrťnalien. Elle a ťgalement de notables effets centraux. Dans ce cadre, elle induit trŤs rapidement une forte sensation de soif. Elle pourrait par ailleurs stimuler les productions d'ACTH et d'ADH hypophysaires ; ces derniers effets ne sont cependant pas encore trŤs clairement ťtablis. L'angiotensine III paraÓt aussi efficace que l'angiotensine II ŗ stimuler la sťcrťtion d'aldostťrone. Son effet vasoconstricteur et ses effets centraux paraissent par contre nettement moindre, voire physiologiquement non significatifs. Les activitťs relatives des angiotensines II et III sont variables d'espŤce ŗ espŤce. Chez l'homme, l'angiotensine III n'interviendrait que pour quelque 10 % dans l'activitť angiotensine totale alors que chez le rat elle serait responsable de prŤs de 60 % de cette activitť.
Chez les poissons, oý l'aldostťrone n'intervient pas dans la balance hydrominťrale, l'angiotensine II provoque la sťcrťtion de cortisol. Chez les amphibiens, les reptiles et les oiseaux oý cortisol et aldostťrone paraissent pouvoir intervenir ensemble, l'angiotensine induit la sťcrťtion des deux hormones.
Figure 9-12BC : Le systŤme rťnine - angiotensine. Effets des angiotensines (B) et structure des angiotensines II chez les vertťbrťs (C). AI - AII - AIII : angiotensines 1, 2 et 3.
d. le facteur natriurťtique
auriculaire
Un autre facteur pouvant intervenir dans le contrŰle de la production
d'aldostťrone est le facteur natriurťtique auriculaire ou atrial
natriurťtique (ANF, cfr. 2.2.7). L'ANF parait en effet
inhiber directement la libťration d'aldostťrone. Il inhibe ťgalement la
sťcrťtion de rťnine, ce qui pourrait rendre compte d'un effet indirect sur la
production d'aldostťrone mais aussi de la diminution de la rťsistance
vasculaire pťriphťrique et donc de la diminution de la pression artťrielle
qu'il provoque. L'ANF induit par contre spťcifiquement une vasoconstriction des
artťrioles effťrentes du glomťrule sans provoquer de modification
concomitante du dťbit d'arrivťe. Il s'ensuit une augmentation de
l'ultrafiltration qui peut Ítre importante. Chez l'homme, le dťbit urinaire
peut ainsi Ítre multipliť par 20 et l'excrťtion de sodium par 30. Ces effets
atteignent un maximum aprŤs 5 ŗ 10 minutes et ne durent en gťnťral pas plus
de 20-25 minutes. L'ANF paraÓt par ailleurs avoir des effets centraux,
antagonistes de ceux de l'angiotensine II, inhibant notamment la sensation de
soif.
e. autres hormones
On sait maintenant depuis prŤs de 40 ans que la prolactine, hormone
ťtroitement associťe aux fonctions de reproduction chez les mammifŤres, a des
effets marquťs sur l'osmorťgulation chez les tťlťostťens. Chez les poissons
d'eau douce, la PRL intervient dans le contrŰle de la permťabilitť ŗ l'eau
et aux ions des tťguments extťrieurs. Elle semble par ailleurs stimuler le
pompage actif branchial et rťnal de NaCl.
La PRL a ťgalement des effets osmotiques marquťs entraÓnant chez diffťrents batraciens et en particulier les tritons, un mouvement du milieu terrestre vers le milieu aquatique oý ils s'accouplent et se reproduisent ("water - drive - effect"). Cet effet est si net qu'il a constituť pendant longtemps un test biologique de mise en ťvidence de la prolactine. Son mťcanisme interne reste peu clair ŗ l'heure actuelle.