3.4.1. Les systèmes néphridiens
3.4.2. Les tubes de Malpighi
Comme nous l'avons déjà signalé précédemment, tous les organes ne participent pas à l'osmorégulation des milieux extratissulaires (cfr. figure 1-9). Seront essentiellement concernés: 1) Chez toutes les espèces: le tube digestif impliqué dans les entrées et les sorties d'eau et de NaCl. Les systèmes rénaux, impliqués dans les sorties d'eau et de NaCl. Les téguments extérieurs impliqués soit dans des entrées soit dans des sorties d'eau et/ou de NaCl. 2) Chez certaines espèces des structures particulières vont également être impliquées. Il s'agit des branchies chez les crustacés et les poissons. Ces organes vont être impliqués dans des mouvements entrants ou sortants d'eau et d'ions. Il s'agit par ailleurs des organes dits extra-rénaux, impliqués dans la sécrétion d'ions Na+et Cl-: glandes nasales, linguales ou sub-linguales de certains oiseaux et reptiles, glandes rectales des sélaciens, papilles annales des larves de moustiques, etc... L'étude des mouvements d'ions et d'eau au niveau de ces structures particulières de même qu'au niveau des branchies ou de la peau sort du cadre d'une approche générale de l'osmorégulation. Elle relève plutôt d'une approche spécialisée écophysiologique, envisageant les spécificités de ces structures correspondant à des adaptations à des biotopes particuliers. Cette étude sera abordée au chapitre 11.
Le tractus digestif sera impliqué chez toutes les espèces sans exception dans des entrées et des sorties d'eau et de NaCl.
La part prise par cet organe dans ces phénomènes va varier en importance suivant les espèces considérées. Les sorties sont normalement de peu d'importance, l'essentiel de l'eau et des ions étant en effet absorbés au niveau intestinal. Les entrées peuvent aller de minimes à essentielles. De peu d'importances voir négligeables chez les poissons d'eau douce par exemple elles deviendront essentielles chez tous les vertébrés terrestres (cfr. ci-avant).
L'entrée d'eau et d'ions au niveau buccal relève essentiellement de la morphologie dite fonctionnelle et ne sera pas considérée ici. Le problème de la mesure de la quantité d'eau ingérée peut être résolu de façon simple et directe pour les animaux terrestres, il est d'approche moins aisée chez les animaux aquatiques. Chez ces derniers, on fait appel à une technique indirecte mettant en œuvre une substance qui n'est pas absorbée au niveau des épithélia, notamment du tractus digestif et qui ne peut être qu'ingérée par voie buccale. Il s'agit à l'heure actuelle de plus en plus couramment de l'inuline, un polymère de glucose dont la concentration est facilement mesurée par une réaction colorimétrique. La quantité d'inuline trouvée dans le tractus digestif d'un animal après un temps donné sera évidemment fonction de la concentration de la substance dans le milieu extérieur et du flux entrant d'eau par voie buccale. Chez les poissons d'eau douce, les quantités d'eau bues varient en général de 1 à 5 ml/100g/jour. Chez les poissons marins, elles varient de 10 à 30 ml/100g/jour. Chez les animaux terrestres, les quantités d'eau ingérées sont en général faibles et éminemment variables, dépendant strictement des conditions extérieures. Elles varient en gros de quelque 0,5 à 6 ml/100g/ jour. De nombreuses espèces désertiques ne boivent pour ainsi dire pas, la fréquence des accès à l'eau étant très limitée. Chez ces espèces la balance hydrique est assurée par l'eau contenue dans la nourriture (eau libre + eau métabolique).
L'essentiel de l'eau et des ions ingérés vont passer dans le sang au niveau intestinal. Chez bon nombre d'espèces, une partie importante de cette absorption a lieu dans les portions terminales de l'intestin. Dans certains cas (insectes, oiseaux), l'intestin aboutit avec les conduits urinaires dans un rectum au niveau duquel peut s'effectuer une absorption importante d'eau et d'ions. Cette absorption est en général très efficace, les fèces produites ne contenant que peu d'eau. Les mouvements d'eau sont osmotiques et suivent un gradient d'hyperosmolarité locale généré par un transport actif d'ions: Na+ essentiellement dans la plupart des cas, K+ dans le rectum des insectes. Ce système est illustré de façon schématique dans la figure 1-13. Les mécanismes particuliers pouvant rendre compte des mouvements actifs transépithéliaux seront envisagés de façon détaillée dans le livre II, traitant de la physiologie cellulaire.
Figure 1-13: A: Les mouvements d'eau et d'ions au niveau du tractus digestif. B: Chez les insectes, la récupération d'eau au niveau du rectum est due à un transport actif de KCl provenant essentiellement des tubes de Malpighi du système rénal. Na, K: espèces ioniques activement transportées.
Comme nous l'avons déjà signalé, les systèmes rénaux interviennent de façon majeure dans l'osmorégulation du sang chez la plupart des espèces. Les problèmes osmotiques n'étant pas essentiels chez les espèces marines isosmotiques, certaines d'entre elles ont perdu leurs structures rénales. C'est le cas des échinodermes et des urocordés (tuniciers). La situation des deutérostomiens épithélioneuriens [pogonophores, stomochordés (entéropneustes et ptérobranches)] paraît moins claire, certaines poches cœlomiques pouvant déboucher sur l'extérieur et d'autres non. Les reins, lorsqu'ils sont présents, jouent également un rôle essentiel dans l'épuration du sang et dans l'élimination de "déchets" solubles (voir chapitre 6). Cette fonction d'excrétion, tout comme celle d'osmorégulation est directement liée aux importants mouvements d'eau qui ont lieu au niveau rénal.
Les mouvements d'eau au niveau des systèmes rénaux peuvent impliquer deux mécanismes différents: une ultrafiltration sous pression ou une diffusion suivant un gradient osmotique engendré par un transport actif d'ions. Sauf chez les insectes, l'entrée d'eau dans les système rénaux s'effectue par ultrafiltration. Il peut y avoir réabsorption d'une partie de cette eau chez différentes espèces. Cette réabsorption s'effectue toujours par diffusion suivant un gradient osmotique (cfr. figure 1-14). La quantité d'urine produite sera donc le résultat d'une ultrafiltration suivie éventuellement d'une réabsorption partielle. La connaissance des quantités ultrafiltrées et réabsorbées est d'un grand intérêt en physiologie rénale. Elle fait intervenir l'utilisation d'inuline; une molécule facilement dosable qui n'est ni sécrétée, ni métabolisée, ni absorbée (la créatinine est parfois utilisée également). L'inuline est injectée dans le sang puis recherchée dans l'urine primaire. Chez les organismes ou l'eau entre dans le rein par mouvement osmotique suite à transport actif, l'inuline passe très mal et n'est pas retrouvée dans l'urine. Chez les autres par contre, elle passe librement dans l'ultrafiltrat. Connaissant la quantité d'inuline et le volume d'urine recueillis en un temps donné ainsi que la concentration de l'inuline dans le sang après injection, il est aisé de calculer la quantité d'eau ultrafiltrée à l'entrée du système rénal dans le même temps. Chez les vertébrés, on parlera d'un taux de filtration glomérulaire, l'ultrafiltration s'effectuant chez la plupart d'entre eux au niveau d'une structure appelée glomérule de Malpighi (voir plus loin). Connaissant la quantité d'eau ultrafiltrée et la quantité d'urine formée dans un même temps, la quantité d'eau réabsorbée s'obtient par simple soustraction ( figure 1-14).
L'utilisation de cette technique a permis de montrer l'existence d'un mécanisme d'ultrafiltration chez tous les animaux étudiés jusqu'à présent possédant des structures rénales, sauf les insectes. Chez ces derniers, l'urine est formée dans des tubes de Malpighi au niveau desquels l'entrée d'eau s'effectue par mouvement osmotique.
Figure 1-14: Mouvements d'eau au niveau rénal. A: invertébrés (néphridies), vertébrés (néphrons). B: insectes (tubes de Malpighi).
L'ultrafiltration est le résultat d'une différence entre les pressions hydrostatique et oncotique du sang à l'entrée du système rénal d'une part et les pressions hydrostatique et oncotique du filtrat se trouvant dans les conduits du système rénal d'autre part (cfr. physiologie cellulaire: mouvements d'eau). Elle est d'autant plus importante que cette différence est grande. On pourrait par ailleurs imaginer que le taux de filtration puisse être fonction du débit sanguin au niveau des unités de filtration. Ce n'est en fait jamais le cas en conditions normales, le taux de filtration étant toujours largement inférieur au débit sanguin rénal et étant par ailleurs largement indépendant des variations de ce dernier. Chez l'homme par exemple, les unités de filtration, situées dans la zone corticale du rein et qui reçoivent plus de 50 % du débit sanguin rénal total, sont irriguées sous un débit de quelque 500 ml/100g/min soit un débit total de quelque 1500 ml/min ou encore 90 l/h. Le taux de filtration par contre sera d'environ 120 ml/min ou encore 173 l en 24 heures, soit nettement moins. Par ailleurs, la production d'urine étant de 1,5 l par 24 heures, l'eau filtrant au niveau rénal est réabsorbée à près de 99 %. Ces valeurs ne sont bien sur qu'indicatives, les quantités d'eau filtrées et les productions d'urine étant éminemment variables en fonction des conditions dans lesquelles se trouve l'organisme considéré. Elles illustrent néanmoins l'importance des mouvements de liquide au niveau rénal.
On remarquera par ailleurs que les taux de filtration augmentent avec le niveau d'évolution des organismes (tableau 1-7). Ceci est à mettre plus en relation avec la fonction d'épuration des reins qu'avec leur fonction d'osmorégulation.
TABLEAU 1-7
Taux de filtration rénale chez différentes espèces
| Espèce | Filtration | Espèce | Filtration |
| Pieuvre Homard Saumon |
0,04 0,07 0,12 |
Grenouille Poule Mammifère |
0,66 1,84 2 à 6 |
Comme nous l'avons déjà signalé précédemment, tous les vertébrés ainsi que les invertébrés terrestres et d'eau douce sont capables d'effectuer une réabsorption d'eau (et donc de NaCl) au niveau rénal. Les structures anatomiques impliquées dans ce système dans l'échelle animale sont cependant loin d'être homologues. On notera donc ici une convergence de mécanisme physiologique qui illustre bien l'importance des systèmes rénaux dans la régulation osmotique sanguine.
Envisageons brièvement l'organisation de ces structures dans le règne animal.
Chez les invertébrés, les unités de base des système rénaux sont des "cœlomoductes", canalicules d'origine mésodermique qui établissent une relation entre la ou les cavités cœlomiques (vestigiales ou non) et l'extérieur. Chez la plupart d'entre eux, ces structures sont appelées néphridies. Chez les insectes, la fonction des néphridies est remplie par les tubes dits de Malpighi, d'organisation fort différente. On trouve également des tubes de Malpighi comme unités de base du système rénal chez d'autres arthropodes comme les myriapodes. Chez certains insectes, on retrouvera, associés à ces structures, des systèmes de filtration analogues aux néphridies et regroupés en organes: reins céphaliques des thysanoures, glandes coxales des chélicérates. Chez les crustacés, on ne retrouve que ces dernières structures et non les tubes de Malpighi: glandes antennaires ou "vertes" des décapodes. On retrouvera également des structures néphridiennes chez les céphalocordés. Les tuniciers (ou urocordés) n'ont pas de système rénal comme nous l'avons déjà signalé. L'organisation anatomique des systèmes néphridiens en structures rénales est éminemment variable. Nous ne l'envisagerons pas en détail ici; quelques schémas résumant la situation sont présentés dans les figures 1-15 et 1-16.
3.4.1. Les systèmes néphridiens
On trouve deux types de néphridies différentes chez les invertébrés: les
protonéphridies et les métanéphridies.
Les protonéphridies sont des structures tubulaires, le plus souvent ramifiées, ouvertes sur l'extérieur par un néphridiopore (pore néphridien ou excréteur) et fermées à l'autre bout. On trouve ces néphridies essentiellement chez des animaux dépourvus de cœlome, la partie terminale close est enclavée dans un tissu parenchymateux, situation très défavorable à l'ultrafiltration. Dans la cavité bat soit une touffe de cils (cellules à flamme) soit un seul grand cil (solénocyte). Le battement de ces cils paraît engendrer une dépression suffisante pour drainer les liquides extracellulaires par ultrafiltration comme le suggère des expériences réalisées avec de l'inuline. Le dispositif filtrant fait intervenir des structures podocytaires analogues à celles du néphron glomérulaire des vertébrés ( figure 1-20). On trouve des cellules à flamme chez les rotifères, les vers plats et certaines annélides. On trouve des solénocytes chez les autres annélides et l'amphioxus.
Les métanéphridies ne sont jamais ramifiées et sont ouvertes aux deux bouts. Le bout se trouvant dans la cavité cœlomique (néphrostome) est en général terminé par un pavillon cilié. Chez différentes espèces (annélides notamment), les conduits métanéphridiens sont plus ou moins directement associés aux gonoductes du système reproducteur (figure 1-16). Chez les mollusques et les crustacés, elles sont organisées en organes rénaux appelés organes de Bojanus chez certains mollusques et glande verte chez certains crustacés. Chez les mollusques, la cavité rénale est associée à la cavité péricardique. Les deux structures présentent une activité de filtration et on y trouve de véritables pelotons tissulaires aux parois richement irriguées. Ce système rénal aboutit à l'extérieur, au niveau de la cavité paléale, par deux conduits néphridiens au niveau desquels viennent s'aboucher les gonoductes. La physiologie des métanéphridies est tout aussi mal connue que celle des protonéphridies. Tout au plus peut-on dire qu'il y a ultrafiltration à l'entrée et qu'une réabsorption d'eau et/ou de NaCl peut être effectuée dans le canal néphridien chez les espèces terrestres et d'eau douce.
Quelques structures de type protonéphridien et métanéphridien sont représentées dans les figures 1-15 et 1-16.
Figure 1-16: A: Système néphridien d'un vers annélide primitif montrant en haut des protonéphridies et en bas des métanéphridies. C: cœlome, G: gonades, Gd: gonoducte, Mn: métanéphridie, Nst: néphrostome, Pn: protonéphridie, Sol: solénocyte. B: Chez certaines espèces, ici l'annélide syllidae Odontosyllis, le gonoducte (cœlomoducte) est aveugle, formant une poche qui s'abouche au canal néphridien ouvert sur le cœlome par un néphrostome.
3.4.2. Les tubes de Malpighi
Les tubes de Malpighi sont des structures tubulaires s'ouvrant sur le rectum et fermées
à l'autre bout que l'on retrouve chez de nombreux arthropodes (insectes notamment). Une
représentation schématique de ce système est présenté dans la figure 1-17
.
Comme nous l'avons déjà signalé, l'entrée d'eau dans les tubes de Malpighi s'effectue non par ultrafiltration mais par mouvement osmotique d'eau suivant un gradient établi par transport actif de K+. Une réabsorption d'eau et d'ions s'effectue au niveau du rectum. Chez certaines espèces, la réabsorption d'eau est tellement efficace que les résidus excrétés sont solides. Par exemple, chez le vers de farine (larve de Ténébrio molitor), une espèce qui peut vivre sans aucun accès à l'eau, la réabsorption d'eau est optimalisée par une organisation particulière des tubes de Malpighi en un complexe dit "cryptonéphridial". Les tubes, au lieu de baigner dans la cavité générale, sont repliés, la partie terminale s'alignant le long de la paroi du rectum. L'ensemble est entouré d'une épaisse membrane périnéphritique formant une chambre périrectale. Cette membrane paraît imperméable à l'eau et être le siège d'un intense transport de K+. Le liquide de la chambre périrectale peut ainsi atteindre une osmolarité allant jusqu'à 4000 mOsm/l (contre quelque 300 dans l'hémolymphe!), générant un important gradient avec le fluide rectal. L'eau paraît passer essentiellement du rectum vers le fluide périrectal et de là dans les portions terminales des tubes de Malpighi. L'eau retournerait vers l'hémolymphe dans la partie antérieure des tubes qui serait les siège d'un transport de K+ de la lumière du tubule vers le sang (figure 1-17).
Figure 1-17: A: Représentation schématique du système excréteur d'un insecte (Rhodnius sp.): 4 tubes de Malpighi s'étendant très loin dans la cavité générale s'abouchent à l'entrée du rectum, à la fin du tube digestif. B: Sécrétion et réabsorption d'eau et d'ions au niveau des tubes de Malpighi et du rectum chez le criquet. C: Système cryptonéphridien de Ténébrio molitor.
Chez les vertébrés, la structure de base des organes rénaux est le néphron. Système analogue à la néphridie des invertébrés, il met en communication une cavité cœlomique ou ce qui en tient lieu avec le milieu extérieur.
Le néphron comprend deux parties physiologiquement et anatomiquement distinctes. Une partie filtrante qui est soit ouverte par un néphrostome sur la cavité cœlomique soit fermée sur un peloton vasculaire portant le nom de glomérule. Une partie tubulaire qui conduit l'ultrafiltrat vers un uretère collecteur tout en modifiant sa composition par des systèmes de réabsorption et de sécrétion. Nous ne nous intéresserons ici qu'à la réabsorption d'eau et de NaCl. Les problèmes de sécrétion et de réabsorption d'autres molécules de même que le rôle de certains néphrons comme conduits génitaux seront envisagés ultérieurement. Les néphrons ouverts, fonctionnels chez les larves et embryons des anammiotes, ne se retrouvent fonctionnels que chez quelques espèces à l'état adulte: lamproies, amphibiens, néphrons primaires des chondrichthyens. Les autres espèces sont pourvues de néphrons fermés: myxines, téléostéens, reptiles, oiseaux, mammifères , néphrons secondaires des chondrichthyens. Chez certaines espèces de téléostéens marins, le glomérule disparaît secondairement formant des reins dits aglomérulés. C'est le cas chez les baudroies, syngnathes et hippocampes notamment ( figure 1-18). Certains téléostéens d'eau douce montrent par contre un néphrostome ouvert secondairement.
Figure 1-18: Néphrons ouverts, fermés et aglomérulés. A: Ouvert à glomérule intracœlomique. B: Ouvert à glomérule intranéphronique. C: Fermé glomérulé. D: Fermé aglomérulé. AA: artère rénale afférente, Ao: aorte dorsale, CN: canal néphrostomial, Coe: cœlome, D: segment distal du tubule, G: glomérule, I: segment intermédiaire du tubule, N: néphrostome, P: segment proximal du tubule, U: uretère.
La partie tubulaire présente toujours au moins deux segments, un proximal et un distal, sauf chez les téléostéens à reins aglomérulés où ce dernier a disparu. Chez les oiseaux et les mammifères, une troisième partie appelée anse de Henlé apparaît entre les deux premières. Elle va jouer un rôle important dans la réabsorption d'eau. Seuls les oiseaux et les mammifères sont capables de produire une urine hypertonique au sang. Un fait qui doit être mis en rapport avec la présence chez ces espèces d'une anse de Henlé, comme nous le verrons plus loin.
Chez les mammifères, tous les néphrons ont une anse de Henlé. Sa longueur est très variable et peut être mise directement en rapport avec la capacité de concentrer l'urine et donc de réabsorber l'eau (voir plus loin). Chez les oiseaux, on trouve environ 90% de néphrons de type reptilien (sans anse de Henlé) localisés en surface et environ 10% de néphrons de type mammalien (avec anse de Henlé) localisés en profondeur. Ces derniers néphrons permettent la production d'une urine concentrée jusqu'à 2 fois par rapport au sang. Cette capacité de concentration est nettement inférieure à celle de certains mammifères, désertiques notamment, qui peut être supérieure à 20 fois (voir plus loin).
La structure générale des reins est éminemment variable. Chez les amniotes, les reins sont formés par la réunion d'un grand nombre de néphrons : 900 à 7.500 par rein chez les squamates, 15.000 à 100.000 chez les oiseaux, 15.000 à 15 millions chez les mammifères (2.104 chez la souris, 106 chez l'homme, 15.106 chez l'éléphant).
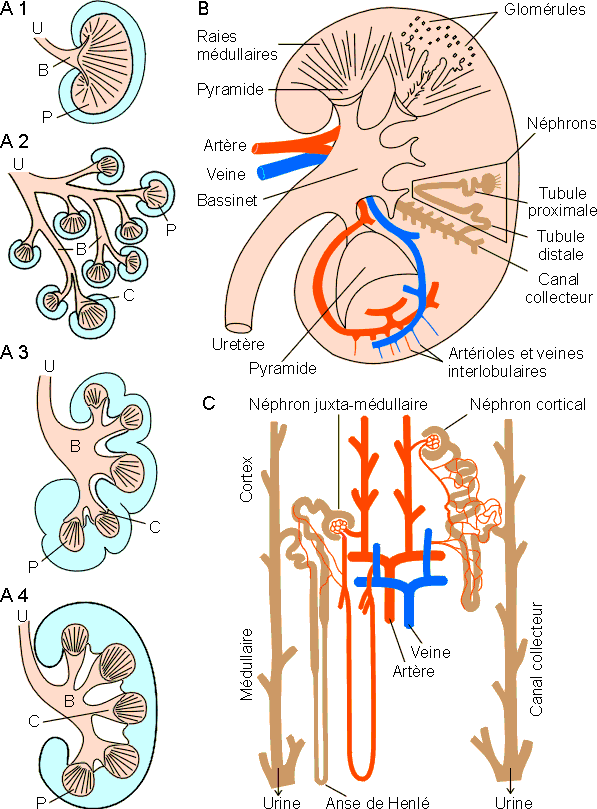
Chez les reptiles et les oiseaux, les glomérules sont centraux, les tubules s'abouchant à des canaux collecteurs périphériques qui se jettent dans l'uretère secondaire. Les glomérules sont de petite taille (± 40 nm de diamètre), peu vascularisés et la filtration glomérulaire est relativement limitée par rapport à ce qu’elle est chez les mammifères. Chez ces derniers par contre, les glomérules sont volumineux (± 200 nm de diamètre) et richement vascularisés ; ce qui permet une filtration beaucoup plus importante. Les glomérules ainsi que les parties contournées des segments proximaux et distaux des tubules sont par ailleurs périphériques, situés dans une partie dite corticale, sombre, d'aspect granuleux. Les anses de Henlé ainsi que les canaux collecteurs et les parties droites des tubes proximaux et distaux sont groupés en une ou plusieurs pyramides dont la base est tournée vers le cortex et le sommet, ou papille, se projette dans la lumière de l'uretère. Les monotrèmes, marsupiaux, rongeurs et primates (sauf l'homme) ont des reins unipyramidaires. Dans ceux ci, l'uretère se dilate en un bassinet où débouchent les tubes collecteurs. Les autres mammifères ont des reins pluripyramidaires dans lesquels le bassinet se ramifie pour se dilater en ampoules secondaires ou calices coiffant chacun une pyramide ( figure 1-19).
Figure 1-19: Structure du rein de mammifère. A: Structure générale chez différentes espèces: rein unipyramidaire (1: lapin) et pluripyramidaires (2, en grappe: cétacé - 3, à lobulation corticale: bœuf - 4, lisse: homme) ; B: bassinet, C: calice, P: pyramide, U: uretère, La partie corticale apparaît en bleu . B: Organisation schématique d'un rein humain. C: Néphrons corticaux (courts) et juxta-médullaires (longs).
Les néphrons sont vascularisés par un double système de capillaires, l'un glomérulaire, l'autre tubulaire. L'irrigation glomérulaire, est toujours artérielle, ne comportant pas de sang veineux. Les capillaires à ce niveau sont fenestrés, permettant une filtration importante (figure 1-20 ; voir aussi chapitre 2, 3.3.3).
L'irrigation tubulaire peut être uniquement artérielle comme chez les cyclostomes où l'irrigation veineuse est absente ou chez les mammifères où elle a disparu secondairement. Chez tous les autres vertébrés, elle est double, artérielle et veineuse ; au sang artériel venant du glomérule s'ajoute un sang veineux venant de la queue et des membres postérieurs.
Une portion importante du système vasculaire tubulaire est le vasa recta qui longe les anses de Henlé et permet la reprise par le système circulatoire de l'eau réabsorbée à ce niveau (figure 1-19).
C |
D E  F  |
Figure 1-20 : Organisation d'un corpuscule rénal (glomérule de Malpighi) de mammifère. L'artériole afférente se divise en un peloton de capillaires dans la chambre glomérulaire, à l'intérieur de la capsule de Bowman (A). Les capillaires (cp) sont ici entourés de cellules particulières, les podocytes (pd), dont les prolongements forment au niveau de la lame basale des capillaires (lb) des fentes de filtration (D, E). La paroi de l'endothélium vasculaire et les prolongements des podocytes forment en définitive un système fenestré facilitant l'ultrafiltration sous pression . Ce système fenestré est bien visible dans l'image F montrant la face interne d'un capillaire en microscopie électronique à balayage.
Chez tous les vertébrés, l'eau ainsi que le NaCl et la plupart des autres solutés entrent dans le système rénal par ultrafiltration. Dans les reins glomérulés, la structure des capillaires (fenestrés) et l'organisation des cellules formant l'unité de filtration, la capsule de Bowman, sont telles qu'elles permettent un passage de liquide important (cfr. figure 1-20 et 5.4.2). De plus, le débit sanguin à ce niveau est l'un des plus important du système circulatoire. Chez l'homme par exemple le débit au niveau des reins peut être estimé à environ 1 à 1,5 litre par minute. L'ultrafiltrat glomérulaire représente lui 150 à 200 litres par jour. Cet ultrafiltrat a la même composition que le plasma sanguin aux protéines près.
La production d'urine chez l'homme étant de 1,5 à 2 l par jour, il est clair que l'essentiel de l'eau, ainsi d'ailleurs que du NaCl et des solutés dits "utiles", est réabsorbé. L'homme réabsorbe ainsi quelque 198 l d'eau et 1,5 à 2 kg de NaCl par jour. La réabsorption d'eau et de NaCl s'effectue au niveau des tubes contournés et de l'anse de Henlé. Elle est liée à un transport actif de Na+ (figure 1-21).
La réabsorption rénale d’eau s’effectue toujours, comme nous l’avons déjà signalé plus haut ( voir 3.3) en suivant un gradient osmotique. Au niveau des tubules contournés, elle est cependant appelée isosmotique. A ce niveau en effet, le gradient osmotique est strictement localisé au niveau des espaces intercellulaires de l’épithélium, l’équilibre osmotique étant maintenu de part et d’autre de celui-ci (cfr. 5.4.2). Ce système est tout à fait différent de celui fonctionnant au niveau de l’anse de Henlé où, comme nous le verrons plus loin, le gradient d’hyperosmolarité s’établit sur une large portion de la médullaire.
Au niveau des tubules contournés, le gradient osmotique local est dû essentiellement à des mouvements actifs de NaCl allant de la face apicale à la face basolatérale de l’épithélium (voir 5.4.2 et 5.4.4 ci-après). Interviennent également à ce niveau les mécanismes de réabsorption de différents solutés organiques comme par exemple le glucose et les acides aminés dont les mouvements sont couplés à des entrées de Na+ (voir 5.4.3). Le passage de ces solutés et du NaCl rompt l’équilibre osmotique qui est immédiatement rétabli par une sortie d’eau. Le système reste donc globalement isosmotique. La réabsorption d’eau est également dans ce cas obligatoire ; ce qui n’est pas le cas dans le système faisant intervenir l’anse de Henlé où la réabsorption d’eau peut être contrôlée par des facteurs hormonaux, indépendamment de l’existence du gradient osmotique. La réabsorption d’eau est d’ailleurs parfois qualifiée dans ce cas de facultative. Comme nous allons le voir, ce sont essentiellement les mouvements de NaCl au niveau de différentes structures qui génèrent au niveau de l’anse un large et important gradient d’hyperosmolarité. L’eau qui est réabsorbée provient essentiellement non pas de l’anse elle-même mais des tubes collecteurs auxquels s’abouchent les néphrons.
L’anse de Henlé se caractérise par sa capacité à générer un important gradient osmotique cortico-papillaire par un système dit d’échange de NaCl à contre courant. Le fonctionnement de ce système peut être mis en rapport avec des caractéristiques particulières des parties ascendante et descendante de l’anse. La partie ascendante est imperméable à l’eau. Elle est de plus le siège d’un intense transport actif de NaCl allant de la lumière vers le sang. Il semble que ce soit le Na+ qui soit l’ion essentiellement transporté. Dans ce schéma, le Cl- suivrait par diffusion en fonction du potentiel transmembranaire (cfr. section 5.4 pour mécanismes intimes). Il se pourrait cependant que du Cl- puisse également être transporté activement. Quoiqu’il en soit, le résultat net, qui est en définitive ce qui nous intéresse ici, est une sortie de NaCl. La partie descendante de l’anse est par contre très perméable à l’eau. Elle est de plus relativement peu perméable au NaCl et ne peut effectuer de transport actif de Na+. Le NaCl pompé spécifiquement dans la branche ascendante va se distribuer par diffusion dans tout l’espace interstitiel de la médullaire y compris dans la branche descendante. Compte tenu de ce que la perméabilité au NaCl de cette branche est relativement basse et qu’elle reçoit un apport continu de liquide isotonique au sang à partir du tube proximal, le NaCl peut y devenir moins concentré que dans le milieu interstitiel environnant lorsque l’activité de pompage dans la branche ascendante est suffisante. Dans ces conditions, une sortie d’eau est induite qui pourrait contribuer à une augmentation de concentration en sel au niveau de la branche descendante. Notons cependant que, comme nous le verrons plus loin, l’essentiel de la sortie d’eau vers le milieu interstitiel s’effectue au niveau des canaux collecteurs.
Le mouvement des ions au niveau de l’anse de Henlé résulte donc en un recyclage du NaCl de la branche postérieure ascendante à la branche antérieure descendante. Le NaCl va ainsi pouvoir s’accumuler dans le fond de l’anse, à la limite jusqu’à saturation des sites de transport se trouvant dans la partie ascendante (figure 1-21).
Figure 1-21: Mouvements d'ions et réabsorption d'eau au niveau des néphrons des mammifères et de certains néphrons des oiseaux.
Cette accumulation et donc le gradient d'hyperosmolarité engendré, sera d'autant plus importante qu'il y a de sites de transport en activité et donc que l'anse de Henlé est longue. Signalons par ailleurs que l'activité de pompage peut être stimulée par l'aldostérone, une hormone du cortex surrénalien (voir chapitre 9, 2.3.2).
Le gradient d'hyperosmolarité ainsi généré peut attirer l'eau qui va diffuser, suivant le gradient osmotique, essentiellement des tubes collecteurs vers l'interstitium médullaire où elle sera reprise par le vasa recta, système de capillaires longeant l'anse de Henlé (figure 1-22). Ces capillaires étant très perméables aux solutés de faibles poids moléculaires, la concentration en NaCl y sera la même que dans l'interstitium médullaire. Ces capillaires sont cependant imperméables aux protéines sanguines qui vont générer une certaine pression oncotique responsable d’un rappel d'eau (voir plus loin section 5.4.2 et aussi chapitre 2, 3.3.2). L’eau passera ainsi en définitive des canaux collecteurs des reins au système circulatoire. Signalons ici que la perméabilité à l'eau des tubes collecteurs est par ailleurs sous la dépendance de l'hormone antidiurétique, ADH (voir chapitre 9, 2.3.2). La réabsorption d’eau à partir des canaux collecteurs est dès lors parfois qualifiée de facultative, par opposition au caractère obligatoire qu’elle présente au niveau des tubules rénaux.
 |
Figure 1-22 : Gradients osmotiques (dégradés colorés), mouvements d'ions ( |
L’intérêt majeur du système de récupération d’eau impliquant l’anse de Henlé est de pouvoir générer un gradient d’hyperosmolarité très important, permettant donc une récupération d’eau de solutions elles-mêmes assez concentrées et dès lors, la production d’une urine également assez concentrée. A la pointe de l'anse de Henlé, dans la région papillaire, l'osmolarité du liquide tubulaire peut atteindre chez l'homme 1400 à 1500 mOsm/l contre ± 300 mOsm/l pour le sang, ce qui représente un pouvoir concentrateur d'environ 5 fois. Chez certaines espèces désertiques, cette capacité de concentration est nettement supérieure, pouvant donner des rapports de concentration urine/plasma allant jusqu'à 31 (tableau 1-8). On remarque une corrélation très nette entre l'épaisseur de la médullaire rénale (la longueur des anses de Henlé) et le pouvoir concentrateur du rein. Un castor, animal n'ayant que peu de problèmes de disponibilité d'eau, aura essentiellement des néphrons courts, corticaux, alors qu'un rat kangourou, espèce désertique, aura des néphrons avec anse de Henlé très longue. Cette relation s’explique évidemment aisément en considérant que le nombre de sites de transport de NaCl se trouvant dans la partie ascendante de l’anse est d’autant plus élevé que celle-ci est longue.
Comme nous l’avons signalé plus haut, la capacité de réabsorption des reins d’oiseaux et de reptiles est relativement faible. L’anse de Henlé n’existe en effet pas au niveau des néphrons des reptiles et les néphrons avec anse ne sont que peu nombreux au niveau des reins des oiseaux. Ces derniers, ainsi que de nombreux reptiles, sont cependant parfaitement capables de produire des déjections parfois très concentrées. En fait, les urines peu concentrées de ces espèces aboutissent avec les fès dans un cloaque au niveau duquel peut encore s’effectuer une réabsorption d’eau. Les mécanismes intimes de ce processus reste à l’heure actuelle très mal connu et ne seront donc pas envisagés ici.
TABLEAU 1-8
Capacité de concentration du rein chez différents mammifères dont les accès
à l'eau sont de plus en plus limités, du castor au rat kangourou.
| Animal | Urine (mOsm/l) | Rapport urine/plasma |
| Castor | 520 | 1,7 |
| Porc | 1100 | 3,6 |
| Homme | 1500 | 5,0 |
| Rat | 3000 | 10,0 |
| Chat | 3100 | 10,3 |
| Rat Kangourou | 5500 | 18,3 |
| Rat des sables | 6300 | 21,0 |
| Souris sauteuse | 9400 | 31,3 |